Анатомия земноводных (Gugmkbnx [ybukfk;ud])

Анатомия земноводных — совокупность сведений о строении тела земноводных.
Земноводные являются первым классом наземных позвоночных животных, из-за чего в их организации имеются как примитивные черты, сохранившиеся от рыбообразных предков, так и прогрессивные, связанные с переходом к наземно-воздушной среде. Покинув водную среду, амфибии выработали принципиально иной способ передвижения, основанный на работе рычажных конечностей. Необходимость в независимых от туловища движений головы и прикрепление пояса задних конечностей к позвоночнику привели к выделению в нём двух новых отделов — шейного и крестцового. Переход к воздушному и отказ от жаберного дыхания потребовали перестройки черепа, а приобретение лёгких привело к возникновению второго круга кровообращения и разделению предсердия на два. Сформировался новый отдел органа слуха — среднее ухо с барабанной перепонкой и стремечком. В нервной системе произошло разделение переднего мозга сквозной щелью на два полушария[1].
Вместе с тем, земноводные сохранили прочную связь с водой. Их индивидуальное развитие как правило протекает со стадией водной личинки, которая становится взрослой особью в ходе метаморфоза. Кожа амфибий лишена чешуи, и потому проницаема для воды и газов, что позволяет осуществлять кожное дыхание, но заставляет держаться возле мест с высокой влажностью[1].
Покровы
[править | править код]
Все земноводные имеют гладкую и тонкую кожу, сравнительно легко проницаемую для жидкостей и газов. Через неё происходит газообмен, что позволяет взрослым амфибиям дышать, не поднимаясь на поверхность воды, а также впадать в спячку на дне водоёмов. Как и других позвоночных животных, в коже земноводных выделяется многослойный эпидермис и собственно кожа (дерма, или кориум). Чтобы увлажнять тонкую и нежную кожу, у земноводных развились слизистые железы, в основном на голове, спине и хвосте. Кроме того, у большинства видов амфибий имеются зернистые железы, выделяющие неприятные на вкус или ядовитые вещества[2].
Эпидермис
[править | править код]Эпидермис состоит из 2—3 слоёв клеток у личинок и 5—7 слоёв у взрослых особей. Клетки наиболее глубокого базального слоя непрерывно делятся и обеспечивают возобновление кожи. У большинства взрослых земноводных в ходе метаморфоза появляется роговой слой (как правило однослойный), клетки которого уплощаются и ороговевают. Он обеспечивает защиту от механических повреждений[3]. Переход от базального слоя к роговому у земноводных постепенный, без резкой границы. Между ними могут выделяться промежуточные слои, но они не всегда чётко выражены[4]. Так, некоторые учёные признают наличие лишь базального и рогового слоёв[5][3], в то время как другие выделяют также зернистый и шиповатый[6]. Помимо кератиноцитов, в эпидермисе также присутствуют бокаловидные клетки, клетки Меркеля и мигрирующие сюда макрофаги, лимфоциты и меланоциты[6]. Эпидермис лежит на базальной мембраной, отделяющей его от дермы и состоящей из коллагеновых волокон[5].
Роговой слой периодически обновляется в процессе линьки, контролируемой гипофизом и щитовидной железой. Наружная часть кожи сбрасывается в основном целиком, в отличие от млекопитающих и птиц, у которых она отслаивается хлопьями. Отслоившуюся кожу земноводные часто поедают[7].
Производные эпидермиса достаточно разнообразны. У прыгуний, веслоногов, квакш, свистунов и древолазов на кончиках пальцев имеются прикрепительные диски. У самцов многих бесхвостых и хвостатых в период размножения на передних лапах образуются брачные мозоли, помогающие удерживать самок во время амплексуса и икрометания[6]. У некоторых представителей, например жаб, на коже имеются местные утолщения (часто называемые бородавками).
Дерма
[править | править код]Дерма состоит из двух слоёв: внешнего рыхлого и внутреннего компактного[5][8]. Рыхлый слой состоит из рыхлой волокнистой соединительной ткани из переплетённых коллагеновых и эластических волокон, желёз, нервов, кровеносных сосудов и вертикально ориентированных гладкомышечных волокон. Более глубокий компактный слой состоит из плотно сложенных коллагеновых волокон. У некоторых бесхвостых (в основном наземных) описан также слой Эберта-Кащенко, расположенный между рыхлым и компактным слоями. Он богат кальцием и полисахаридами (преимущественно хондроитинсульфатом) и, предположительно, накапливает в себе воду[9]. У хвостатых и безногих компактный слой дермы плавно переходит в соединительную ткань, окружающую мышцы и кости. У бесхвостых кожа отделена от тела лимфатическими мешками и сохраняет контакт лишь в некоторых местах. Он может осуществляться прозрачными листками соединительной ткани (лимфатическими септами), пучками соединительнотканных волокон, гладкими или поперечнополосатыми мышцами, а также остеодермами, которые могут быть как васкуляризироваными (например у тигровой сумчатой квакши и двухцветной филломедузы), так и бессосодустыми (у Pelobatrachus nasutus и Craugastor augusti). У жаб и некоторых квакш может происходить окостенение дермы над элементами крыши черепа[5].
У безногих земноводных в карманах дермы залегают чешуйки, состоящие из уплощённых клеток толщиной 4—5 мкм, которые откладывают на своей поверхности плотно упакованные коллагеновые волокна, минерализованные кремнием, цинком, кальцием и фосфором[9].
Кожные железы
[править | править код]В дерму погружено большое количество альвеолярных желёз, имеющих эктодермальное происхождение. Они делятся на слизистые, выделяющие мукополисахариды и зернистые, выделяющие различные ароматические амины и полипептиды, некоторые из которых являются токсинами. Слизистые железы обычно меньше зернистых и полностью погружены в рыхлый слой дермы. У безногих они обладают хорошо выраженными миоэпителиальными клетками, однако у некоторых бесхвостых и хвостатых их нет. Зернистые железы крупнее и своими основаниями могут погружаться в компактный слой дермы. У них имеются миоэпителиальные клетки, иногда двух типов, а у некоторых древолазов зернистые железы окружены также меланофорами[5].
Распределение желёз на теле неоднородно. Как правило, слизистые железы более многочисленны на спинной стороне тела. Зернистые железы часто собираются в группы на определённых участках тела, например на голове и шее у многих бесхвостых и некоторых хвостатых и на спинной стороне хвоста у других хвостатых. Некоторые зернистые железы собираются в хорошо различимые внешне структуры, такие как бородавки на спине и паротиды жаб и некоторых саламандр, поясничные железы некоторых родов свистунов, железы на голенях некоторых австралийских и настоящих жаб, а также складки на спине настоящих лягушек. Выделения таких желёз часто имеют защитную функцию. Кроме слизистых и зернистых желёз, у некоторых амфибий известны ещё три вида желез: железы в кожных складках червяг, предположительно секретирующих материал чешуек, липидные железы филломедуз, предохраняющие их кожу от высыхания, и клеевые железы на груди узкоротов, обеспечивающих приклеивание самцов к самкам во время амплексуса[5].
Хроматофоры
[править | править код]
Цвет кожи земноводных определяется тремя типами пигментных клеток, называемых хроматофорами: меланофорами, содержащими эумеланин, который даёт чёрную, коричневую или красную окраску; иридофорами, или гуанофорами, содержащих пурины (гуанин, гипоксантин, аденин) и отражающих свет определённых волн; и ксантофорами, или эритрофорами, содержащих птеридин или каротиноиды, дающих жёлтый, оранжевый или красный цвета. Меланофоры могут присутствовать в эпидермисе у личинок, у которых они являются главными пигментными клетками. Они выглядят как тонкие вытянутые клетки с ветвящимися отростками, залегающими между соседними мальпигиевыми клетками, содержащими рецепторы к меланину. В ходе метаморфоза эпидермальные меланофоры редуцируются, и у взрослых земноводных пигментные клетки располагаются в дерме, образуя слои. Наиболее поверхностное положение занимают ксантофоры, располагаясь сразу под базальной мембраной, хотя у некоторых земноводных, например зеленоватого тритона, они могут присутствовать в эпидермисе. Под ксантофорами располагаются белые или серебристые иридофоры. Вместе с ксантофорами они создают яркие цвета. В них содержатся специальные отражающие свет органеллы, лежащие параллельными стопками. Как правило, иридофоры расположены в один слой, но у хватающих лягушек, обитающих в очень засушливых условиях, в коже имеется 3—5 слоёв иридофоров, что увеличивает отражение света и снижает испарение воды с поверхности тела. Глубже всех расположены меланофоры. Своими отростками они обхватывают иридофоры, отделяя их ксантофоров. У некоторых представителей подсемейств Phyllomedusinae и Pelodryadinae меланофоры помимо меланина содержат красный пигмент птерородин[англ.] (димер птеридина), не встречающийся у других позвоночных[5].
Изменение цвета, наблюдаемое у многих видов, инициируется гормонами, выделяемыми гипофизом. В отличие от костистых рыб, нервная система не контролирует пигментные клетки напрямую, в результате чего изменение цвета происходит медленнее, чем у рыб. Ярко окрашенная кожа обычно указывает на то, что этот вид ядовит, и является предупреждающим знаком для хищников[10][11].
Скелет и мускулатура
[править | править код]

Скелет земноводных соответствует общему плану строения наземных позвоночных. Все амфибии, за исключением безногих и нескольких видов хвостатых, имеют две пары конечностей. Кости полые и лёгкие[12].
Череп
[править | править код]Череп земноводных представляет собой единую малоподвижную структуру[13]. Из-за редукции покровных окостенений, у современных земноводных череп во взрослом состоянии сохраняет много хряща[14]. Сам череп широкий и уплощённый, что повышает объём ротоглоточной полости и увеличивает эффективность всасывания воздуха в неё. Осевой череп платибазальный, то есть имеет широкое основание, позволяющее мозгу располагаться между глазницами[13]. В затылочном отделе окостенению подвергаются лишь парные боковые затылочные кости, каждая из которых несёт по одному затылочному мыщелку, соединяющиеся с шейным позвонком. В слуховой области находятся парные переднеушные кости, а в глазничной — парные глазоклиновидные у хвостатых амфибий или сросшаяся кольцевидная клиновиднообонятельная кость у бесхвостых. Крыша черепа образована парными теменными и лобными костями, которые срастаются в парные лобнотеменные у бесхвостых. Впереди от них расположена пара носовых костей и (у хвостатых) 1—2 пары предлобных. В слуховой области образуется чешуйчатая кость, также имеющая покровное происхождение[14]. Дно черепа покрыто крупным крестовидным парасфеноидом[13] Перед ним лежат парные нёбные кости и сошники. У хвостатых они сливаются в нёбносошниковые кости. На сошниках или нёбносошниковых костях сидят мелкие зубы[14].

Висцеральный череп образован нёбноквадратным хрящом, который сохраняется в течение всей жизни. Передним и задним концами он прирастает к черепной коробке, что обеспечивает аутостилию. К нему же прилегают предчелюстная и верхнечелюстная кости, на которых сидят мелкие зубы. Задний участок нёбноквадратного хряща покрыт сверху квадратно-скуловой и чешуйчатой костями, а снизу — квадратной. У некоторых хвостатых земноводных он окостеневает, образуя квадратную кость[14].
Нижняя челюсть образована меккелевым хрящом, который окостеневает лишь передним концом, образуя подбородочно-челюстные кости. За ними лежат покровные зубные кости, которые у современных земноводных не имеют зубов. Заднюю часть хряща обрастает длинная угловая кость и несколько мелких добавочных косточек. Суставным отростком меккелев хрящ сочленяется с задним концом нёбно-квадратного хряща, образуя челюстной сустав[14].
В связи с переходом от гиостилии к аутостилии произошла редукция подъязычной дуги. Её верхний элемент — гиомандибуляре — преобразовался в слуховую косточку (стремечко), а нижний — гиоид — вместе с формирующимися у личинок жаберными дугами формируют подъязычную пластинку, которая составляет твёрдое дно ротоглоточной полости. Она несёт две пары крупных рожков[13].
-
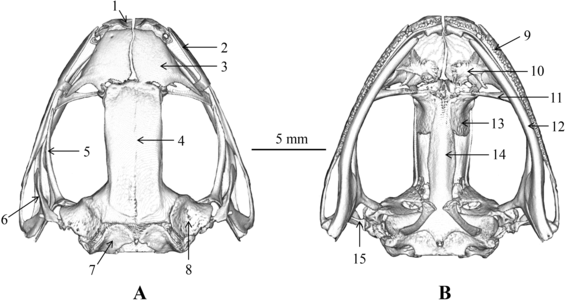 Череп лягушки Nidirana leishanensis (A) Сверху. (B) Снизу. 1 — предчелюстная кость, 2 — верхнечелюстная кость, 3 — носовая кость, 4 — лобнотеменная кость, 5 — крыловидная кость, 6 — чешуйчатая кость, 7 — боковая затылочная кость, 8 — переднеушная кость, 9 — зубы верхней челюсти, 10 — предсошник, 11 — нёбная кость, 12 — нижняя челюсть, 13 — клиновиднообонятельная кость, 14 — парасфеноид, 15 — стремечко
Череп лягушки Nidirana leishanensis (A) Сверху. (B) Снизу. 1 — предчелюстная кость, 2 — верхнечелюстная кость, 3 — носовая кость, 4 — лобнотеменная кость, 5 — крыловидная кость, 6 — чешуйчатая кость, 7 — боковая затылочная кость, 8 — переднеушная кость, 9 — зубы верхней челюсти, 10 — предсошник, 11 — нёбная кость, 12 — нижняя челюсть, 13 — клиновиднообонятельная кость, 14 — парасфеноид, 15 — стремечко -
 Череп большеногой саламандры. (a) — сверху, (b) — снизу. fr = лобная кость, mx = верхнечелюстная кость, n = носовая кость, nlc = носослёзный канал, oc = слуховая капсула, opl = opercular plate, osph = глазоклиновидная кость, pa = теменная кость, pat = отросток теменной кости, pmx = предчелюстная кость, prf = предлобная кость, psph = парасфеноид, ptrp = отросток крыловидной кости, q = квадратная кость, sm = септомаксилла, sq = чешуйчатая кость, v = сошник, vpop = предглазничный отросток сошника, pvtp = задний ряд сошниковых зубов
Череп большеногой саламандры. (a) — сверху, (b) — снизу. fr = лобная кость, mx = верхнечелюстная кость, n = носовая кость, nlc = носослёзный канал, oc = слуховая капсула, opl = opercular plate, osph = глазоклиновидная кость, pa = теменная кость, pat = отросток теменной кости, pmx = предчелюстная кость, prf = предлобная кость, psph = парасфеноид, ptrp = отросток крыловидной кости, q = квадратная кость, sm = септомаксилла, sq = чешуйчатая кость, v = сошник, vpop = предглазничный отросток сошника, pvtp = задний ряд сошниковых зубов


Осевой скелет
[править | править код]
В позвоночнике выделяют 4 отдела: шейный, туловищный, крестцовый и хвостовой. Шейный отдел состоит только из одного позвонка, несущего две небольшие суставные впадины для сочленения с затылочными мыщелками[13]. Из-за этого голова земноводных может совершать лишь движения вверх-вниз, в отличие от млекопитающих и птиц, имеющих зубовидный отросток второго шейного позвонка, по которому проходит ось вращения[15]. На туловищных позвонках хорошо развиты поперечные отростки, к которым причленяются короткие рёбра. При этом у большинства бесхвостых, кроме наиболее примитивных семейств (лейопельмы, круглоязычные, жерлянки и пиповые, у которых они присутствуют на первых трёх туловищных позвонках[16]) они редуцированы и сливаются с поперечными отростками. Крестцовый отдел, как и шейный, состоит из единственного позвонка. К концам его поперечных отростков причленяются подвздошные кости тазового пояса[14]. В связи с редукцией пояса задних конечностей, у безногих земноводных крестцовый отдел не выделяют[16]. Хвостовые позвонки хвостатых амфибий несут нижние дуги, образующие гемальный канал. У бесхвостых все хвостовые позвонки сливаются в единую кость — уростиль[14].
У наиболее примитивных земноводных (безногих, некоторых хвостатых и бесхвостых) позвонки амфицельные, то есть с двояковогнутыми телами. У большинства хвостатых и некоторых бесхвостых позвонки опистоцельные (тела выпуклые спереди и вогнутые сзади), а остальные имеют процельные позвонки (тела вогнуты спереди и выпуклы сзади)[14]. Число позвонков сильно варьирует. Наиболее укорочен позвоночник бесхвостых, состоящий из 1 шейного, 4—7 туловищных и одного крестцового позвонков. Позвоночник безногих может включать 60—285 позвонков[16]. У хвостатых 13—62 туловищных и 22—36 хвостовых позвонков[14].
Скелет конечностей
[править | править код]
Парные конечности земноводных, в отличие от парных плавников рыб, имеют только внутренний скелет и построены по принципу системы рычагов, соединённых суставами. Связь конечностей с туловищем осуществляется поясами конечностей, устроенными иначе, чем у рыб. Так, в поясе передних конечностей количество элементов сокращается, а сам он теряет связь с черепом, в то время как пояс задних конечностей соединяется с крестцовым позвонком и увеличивается в размерах[17]. Обе пары конечностей устроены по одной схеме, общей для всех наземных позвоночных. В них выделяют три отдела[18]:
- плечо/бедро — состоит из плечевой или бедренной кости;
- предплечье/голень — состоит из лучевой и локтевой или большой и малой берцовых;
- кисть/стопа — состоит из 3 подотделов:
- запястье/предплюсна — из 9—10 косточек, расположенных в три ряда;
- пясть/плюсна — из пяти удлинённых костей, расположенных веерообразно в один ряд;
- фаланги пальцев — из пяти или меньше рядов, продолжающих кости пясти или плюсны, по нескольку косточек в каждом.
Вместе с тем, у всех современных земноводных на передней конечности развивается не более четырёх пальцев[14]. Кроме того, часто редукции подвергается количество фаланг на пальцах. Так, если у анцестральных земноводных фаланговая формула имела вид 2—3—4—5—4, то для современных амфибий характерны варианты 1—2—3—2 и 2—2—3—3[17].

Плечевой пояс лежит в толще мускулатуры и имеет вид полукольца. Он включает парные лопатки, ключицы и коракоиды, соединённые с грудиной. Коракоид и лопатка образуют суставную впадину, с которой сочленяется головка плеча. К лопатке присоединён широкий надлопаточный хрящ, к которому присоединяются мышцы спины. Перед коракоидом лежит хрящевой прокоракоид, имеющий палочковидную форму и прикрываемый снизу ключицей. Костная грудина имеет хрящевое расширение на заднем конце. При этом у хвостатых амфибий в плечевом поясе сохраняются лишь лопатка, прокаракоид и коракоид, которые сливаются и окостеневают лишь в районе суставной впадины, а также грудина, состоящая из хряща. У безногих плечевой пояс редуцировался в связи с утратой конечностей[17].
Передние конечности большинства хвостатых земноводных по своему строению незначительно откланяются от общей схемы. Отличаться может лишь количество пальцев, которое у большинства представителей равно четырём, а у протеев — трём[14]. Кости плеча и предплечья обычно состоят из хряща на концах, а элементы кисти или остаются полностью хрящевыми, или окостеневают лишь в центральной части. У бесхвостых земноводных скелет свободных конечностей окостеневает полностью[17] и претерпевает более значительные структурные изменения, которые обусловлены адаптациями к движению прыжками. Локтевая и лучевая кости сливаются в единую кость предплечья. Уменьшается число косточек запястья, а также появляются 1—2 добавочные косточки, которые являются рудиментами предпальца[14].

Тазовый пояс состоит из парных подвздошных, седалищных и лобковых хрящей, которые, срастаясь, образуют вертлужную впадину. Подвздошная кость расположена расположена дорсально и сочленяется с поперечными отростками крестцового позвонка. Седалищная кость и лобковый хрящ расположены вентрально и вместе образуют хрящевую пластинку[17]. У хвостатых земноводных к ней спереди причленяется раздвоенный на конце надлобковый хрящ, который поддерживает стенку брюшной полости. У бесхвостых подвздошные кости заметно удлинены, а седалищные кости срастаются между собой[19].
Задние конечности хвостатых земноводных окостеневают сходным образом с передними. Как правило, на задних лапах у них имеется пять пальцев с фаланговой формулой 1-2-3-3-2, хотя у некоторых представителей, например четырёхпалой саламандры, наблюдается редукция пятого пальца. У бесхвостых задние конечности увеличиваются относительно передних. Кости голени срастаются в единую кость[17]. Две кости проксимального ряда предплюсны удлинены, образуя добавочный рычаг, в то время как дистальный ряд редуцируется до 2-3 косточек, к которым примыкает две косточки рудимента предпальца. Эти косточки составляют скелет пяточного бугра[14]. За исключением некоторых видов, на задних лапах бесхвостых имеется пять пальцев с фаланговой формулой 2-2-3-4-3[17].
Мускулатура
[править | править код]Мышечная система амфибий под влиянием перехода к наземному образу жизни претерпела значительные изменения. Значительно дифференцируется осевая мускулатура, хотя часть туловищной мускулатуры образована миомерами, разделёнными миосептами, сохраняя первичное метамерное строение[20]. Появляется развитая и сложно организованная мускулатура конечностей, основанная на действии мышц-сгибателей и мышц-разгибателей. Специализируется мускулатура ротовой полости, в том числе жевательные мышцы, мышцы языка и дна ротовой полости. Они играют важную роль не только в захвате и удержании пищи, но и обеспечивают акт дыхания[14].
Большой специализации достигает мускулатура бесхвостых амфибий, что обусловлено их адаптацией к передвижению прыжками. В связи с редукцией хвоста и укорочением туловища, их осевая мускулатура слабо сегментирована. Вместе с тем, большее развитие получили мышцы, обеспечивающие вертикальное изгибание позвоночника, а также мышцы конечностей, обеспечивающие прыжки[21].
Дыхательная система
[править | править код]Земноводные используют для газообмена более разнообразные органы, чем любой другой класс позвоночных[22]. В дыхании могут участвовать: жабры (как наружные, так и внутренние), кожа, слизистая оболочка ротоглоточной полости и лёгкие[23].
Жабры
[править | править код]
У личинок земноводных на жаберных дугах образуются перистые кожные выросты, покрытые складочками — наружные жабры. Они пронизаны многочисленными капиллярами, через стенки которых происходит газообмен[23]. У личинок хвостатых и безногих амфибий они остаются до метаморфоза, в то время как у бесхвостых они остаются рудиментарными и редуцируются в ходе развития, заменяясь внутренними жабрами[24]. Они омываются водой, поступающей через рот и выводящейся наружу через жаберные щели. Через несколько дней после вылупления у жаберные щели с каждой стороны тела покрываются кожистыми жаберными крышками, которые сзади прирастают к телу. При этом вода выводится наружу через небольшое отверстие с левой стороны тела[23]. Обычно жабры исчезают при метаморфозе, однако у некоторых земноводных, например американских протеев, они сохраняются в течение всей жизни в связи с тем, что они не проходят метаморфоз, оставаясь неотеническими личинками[22].
Лёгкие
[править | править код]_dissection_-_(3).jpg)
В ходе метаморфоза у большинства земноводных появляются лёгкие. Они представляют собой полые мешки, имеющие ячеистую складчатость. Каждое лёгкое открывается самостоятельным отверстием в гортанно-трахейную впадину, которая отграничена гортанными хрящами и открывается щелью в ротоглоточную полость. На стенках камеры расположены голосовые связки, вибрация которых обеспечивает вокализацию. У бесхвостых гортанно-трахейная впадина укорочена, а у хвостатых и безногих она развита и может рассматриваться как зачаток трахеи[23]. В отличие от других земноводных, у пиповых гортанно-трахейная впадина разветвляется на конце, образуя бронхи[24].
Площадь дыхательной поверхности лёгких меньше поверхности тела[23]. Однако у разных видов она отличается. Так, у живущих в холодных ручьях олимпийских амбистом лёгкие небольшие и гладкие внутри, в то время как у карликового сирена они удлинённые, а их внутренняя поверхность благодаря многочисленным складкам, превышает внешнюю в три раза. У съедобной лягушки внутренняя поверхность превышает внешнюю в восемь раз[22]. Высокой изменчивостью обладает и длина капиллярной сети, оплетающей лёгкие. Так, у полуводного гребенчатого тритона отношение длины капилляров кожи к длине капилляров лёгких составляет 3:1, у влаголюбивых чесночниц и жерлянок — 1:1, а у сухопутных жаб — 1:3[23].
В связи с роющим образом жизни у безногих земноводных лёгкие как правило хорошо развиты и имеют высокую складчатость[22]. Например, лёгкие бразильской червяги[англ.] достигают последней задней трети тела, а у водной червяги они доходят до его конца, загибаясь в петли[25]. У многих видов лёгкие развиты несимметрично. Например у Ichthyophis orthoplicatus правое лёгкое в четыре раза больше левого, а у большинства наземных видов, таких как кольчатая червяга, — в 10 раз. В то же время у Hypogephis rostratus левое лёгкое находится в зачаточном состоянии, а у Boulengerula taitanus оно вовсе отсутствует[22].
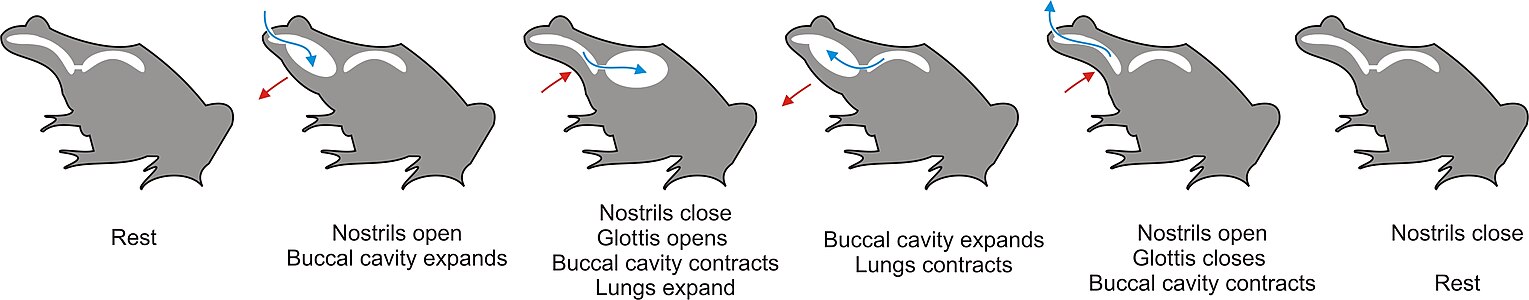
В связи с отсутствием грудной клетки воздух нагнетается в лёгкие движениями дна ротоглоточной полости. Такой тип дыхания называется гулярным и осуществляется следующим образом: при опускании дна воздух насасывается в ротоглоточную полость через открытые ноздри. После закрытия наружных ноздрей открывается гортанная щель, и воздух из лёгких смешивается с атмосферным в ротоглоточной полости. Дно полости поднимается и смешанный воздух нагнетается в лёгкие. После этого ноздри открываются и остатки смешанного воздуха выводятся наружу. В промежутках между дыхательными движениями дно ротовой полости совершает меньшие по амплитуде колебания при открытых ноздрях и закрытой гортани, что позволяет обновлять воздух в ротоглоточной полости, через пронизанную капиллярами слизистую оболочку которой также осуществляется газообмен[23][26].
- Механизм гулярного дыхания
-

Некоторые земноводные полностью утратили лёгкие. Среди них безлёгочные саламандры, дальневосточные тритоны, червяга Айзельта Microcaecilia iwokramae[англ.] и калимантанская барбурула. Газообмен у этих земноводных происходит через кожу[27] или (как у червяг, чья кожа значительно утолщена) через слизистую оболочку ротоглоточной полости[25].
Кожа
[править | править код]
Большинство земноводных способны осуществлять газообмен через кожу как в воде, так и на воздухе. Лёгочное дыхание сохраняется относительно постоянным в течение года, тогда как интенсивность кожного дыхания отчётливо возрастает весной и летом и снижается зимой. Слизистая оболочка полости рта также участвует в газообмене. Чтобы обеспечить достаточное дыхание через кожу, поверхность сильно васкуляризированной кожи должна оставаться влажной, позволяя молекулам кислорода диффундировать с достаточной скоростью. Поскольку концентрация кислорода в воде увеличивается при низких температурах, водные амфибии в этих условиях полагаются в основном на газообмен, осуществляемый через кожу. Они в основном небольших размеров (что означает большое отношение площади поверхности тела к его объёму), но не характеризуются чрезмерно развитой сетью капилляров в коже. У червеобразных саламандр он развит даже слабо. Это компенсируется большим количеством ядерных эритроцитов. У некоторых видов, например тихоокеанской амбистомы дорсальные капилляры вдвое толще вентральных. Во время сезона размножения у самца волосатой лягушки развиваются волосовидные выросты кожи, которые играют роль в функционировании наружных лёгких[22].
Чтобы кислород мог диффундировать через неё с достаточно высокой скоростью, поверхность кожи должна оставаться влажной. Поскольку концентрация кислорода в воде увеличивается как при низких температурах, так и при высоких скоростях потока, водные амфибии в этих ситуациях могут полагаться в первую очередь на кожное дыхание. Титикакский свистун может всю жизнь дышать кожей[12].
Сердечно-сосудистая система
[править | править код]Кровеносная система личинок сохраняет примитивное строение, характерное для рыб. У них имеется один круг кровообращения и двухкамерное сердце, состоящее из одного предсердия и одного желудочка. Кроме того, перед предсердием расположен венозный синус, собирающий венозную кровь, а из желудочка кровь попадает в артериальный конус, который разветвляется на четыре пары приносящих жаберных артерий. Первые три пары распадаются на капилляры в жабрах, а четвёртая впадает в корни спинной аорты. Выносящие жаберные артерии первой жаберной дуги, распадаясь на сонные артерии, снабжают кровью голову. Артерии второй и третьей дуг сливаются в правый и левый корни аорты, которые объединяются в спинную аорту[28].

У большинства взрослых земноводных, в связи с появлением лёгочного дыхания, возникает малый, или лёгочный круг кровообращения. В ходе метаморфоза единое предсердие разделяется на левое и правое, и сердце становится трёхкамерным (хотя у хвостатых и безногих перегородка между предсердиями неполная[29]). К правому предсердию примыкает венозный синус. В него собирается кровь от органов. В левое предсердие от органов дыхания поступает насыщенная кислородом артериальная кровь. Оба предсердия открываются общим отверстием в желудочек, из-за чего кровь смешивается. Однако смешение крови неполное, что обеспечивается мускулистыми выростами внутренних стенок желудочка, которые образуют ряд сообщающихся камер. От правой стороны желудочка, где сконцентрирована более венозная кровь, отходит артериальный конус, снабжённый спиральным клапаном[28] — продольной складкой, распределяющей кровь при сокращении сердца[30]. У некоторых безлёгочных саламандр он отсутствует[31]. В механизме распределения крови во время систолы большое значение имеет пониженное сопротивление капилляров в лёгких, из-за чего кровь из желудочка сначала устремляется в кожно-лёгочные артерии, и лишь при их заполнении — в сонные артерии и дуги аорты[32].
Артериальный конус переходит в луковицу аорты, которая разделяется на две ветви. Каждая из них внутри поделена на три канала: передний — сонный, средний — системный и задний — кожно-лёгочный. Они продолжаются в самостоятельные артериальные дуги: общую сонную артерию, дугу аорты и кожно-лёгочную артерию соответственно. Общая сонная артерия снабжает артериальной кровью голову. Сразу после отделения она разветвляется на наружную (питает подъязычную мускулатуру, глотку и язык) и внутреннюю (снабжает глаза, нёбо и мозг) сонные артерии. Правая и левая дуги аорты огибают с обеих сторон пищевод и сливаются в спинную аорту. Перед слиянием от них отходят пищеводная, затылочно-позвоночная и подключичная артерии. Спинная аорта тянется вдоль позвоночника. От неё отходят артерии, питающие внутренние органы. Кожно-лёгочная артерия распадается на кожную и лёгочную артерии, которые несут кровь к органам дыхания (коже и лёгким соответственно)[30].
Венозная кровь от заднего отдела тела и конечностей собирается бедренными и седалищными венами, которые сливаются в воротные вены почек. Кроме того, от бедренных вен отходят вены, сливающиеся в брюшную вену. Вместе с воротной веной печени, собирающей кровь от кишечника и желудка, она распадается на капилляры в печени, образуя воротную систему печени. Выносящие вены почек сливаются в непарную заднюю полую вену. В неё же впадают вены от половых желёз. Проходя через печень, она принимает кровь от почечных вен и впадает в венозный синус. Насыщенная кислородом кровь от кожи собирается в большую кожную вену. Вместе с плечевой веной, несущей венозную кровь она впадает в подключичную вену. Подключичные вены сливаются с наружными и внутренними ярёмными венами, собирающими кровь от головы, в правую и левую передние полые вены, которые независимо впадают в венозный синус. От лёгких обогащённая кислородом кровь собирается в парные лёгочные вены, впадающие в левое предсердие[28].
В связи с повышением давления в кровеносной системе, большее развитие, чем у рыб, получает лимфатическая система, которая предназначена для возврата тканевой жидкости в кровяное русло. Лимфатические сосуды особенно многочисленны под кожей, где образуют обширные лимфатические полости, что предохраняет кожу от высыхания[32]. У хвостатых и безногих земноводных лимфатические сосуды впадают в вены в многочисленных более или менее метамерно расположенных местах. Тем не менее, такие соединения часто концентрируются около передних и задних кардинальных, ярёмных и задней полой вены. Циркуляция лимфы обеспечивается лимфатическими сердцами — мелкими двухкамерными гладкомышечными структурами, расположенные в местах впадения лимфатических сосудов в вены. У безногих их может быть около сотни в парных рядах, а у бесхвостых и хвостатых они менее многочисленные, но присутствует пара сильно развитых сердец в тазовой области[33].

У земноводных впервые появляется красный костный мозг, обеспечивающий гемопоэз. Другими кроветворными органами являются тимус, лежащий в глубине и по бокам шеи и обеспечивающий пролиферацию лимфоцитов на ранних стадиях развития, селезёнка, которая у большинства хвостатых тянется вдоль кишки, а у остальных представителей расположена более компактно. У амфибий сохраняет свою функцию на протяжение всей жизни. Кроветворную функцию имеют также почки и печень, причём у хвостатых последняя даёт значительную долю лейкоцитов. Кроме того, в стенках ротовой полости и глотки земноводных имеются миндалины[34].
Общее количество крови земноводных составляет 1,2—7,2 % от массы тела[28]. Эритроциты земноводных считаются самыми крупными среди позвоночных. Так, у хвостатых земноводных они могут достигать 40—70 мкм (наиболее крупные у амфиум), при этом у личинок они обычно больше, чем у взрослых. Тем не менее, у бесхвостых амфибий они меньше, чем у хвостатых, имея длину 17,7—26,5 мкм[31]. Эритроциты земноводных овальной формы и в основном имеют ядро[34], однако у безлёгочных саламандр 1—5 % эритроцитов могут быть безъядерными, а у некоторых представителей трибы Bolitoglossini — до 95 %[31][35]. Крупные размеры красных кровяных клеток компенсируются относительно небольшим их числом: от 20 до 730 тысяч в 1 мм³ крови. При этом у личинок этот показатель ниже, чем у взрослых[28]. Тромбоциты имеют форму веретенообразных клеток, которые разрушаются, лишь попадая кровяное русло[34].
Пищеварительная система
[править | править код]Ротоглоточная полость широкая, сверху ограничена почти плоским нёбом[36], образованным дном мозгового черепа. В нём имеются внутренние ноздри — хоаны[37]. Как и вся ротовая полость, нёбо выстлано многослойным плоским ресничным эпителием. На дне ротоглоточной полости находится мускулистый язык, своим основанием укреплённый подъязычным аппаратом, что позволяет ему совершать разнообразные движения[36]. У безногих и хвостатых язык прикрепляется ко дну ротоглоточной полости. У безногих он не способен выдвигаться, и во время ловли добычи остаётся на дне рта. У некоторых хвостатых, например грибоязычных саламандр[англ.], язык может иметь грибоподобную форму, прикрепляясь ко дну «ножкой», что позволяет выбрасывать его на треть длины тела (без хвоста) вперёд и вбок[38]. У бесхвостых он передним концом прикрепляется к нижней челюсти, а его свободный конец при ловле добычи выбрасывается изо рта[39]. У самых крупных представителей длина языка может достигать 7—11 сантиметров[источник не указан 742 дня]. Некоторые земноводные, перешедшие к водному образу жизни, утратили язык, и ловят добычу, засасывая её в рот[38]. В ротоглоточную полость открываются протоки слюнных желёз. В передней части нёба находится крупная межчелюстная железа трубчатого строения. Кроме того, выделяются нёбные железы и мелкие железы на языке. Секрет слюнных желёз у большинства амфибий не содержит пищеварительных ферментов, лишь смачивая пищу. Лишь у некоторых бесхвостых обнаруживается амилаза, участвующая в переваривании крахмала[40].

У примитивных вымерших земноводных зубы сохраняли складчатую структуру дентина, характерную для кистепёрых рыб. В связи с этим базальных амфибий выделяли в группу лабиринтодонтов. У современных земноводных, относящихся к группе беспанцирных, зубы относятся к так называемому педицилярному типу. Между основанием и вершиной таких зубов имеется зона слабого обызвествления, а дентин не имеет складчатости[36]. Как правило зубы земноводных двувершинные, но у некоторых хвостатых и бесхвостых могут встречаться одновершинные или многовершинные зубы. Как правило, зубы не отличаются по форме, однако у некоторых видов хвостатых и безногих развилась гетероднотная зубная система. По способу прикрепления зубы земноводных относятся к плевродонтному (на челюстях) и акродонтному (на нёбе) типам. Для амфибий характерен полифиодонтизм, то есть многократная смена зубов в течение жизни[41]. У всех безногих и большинства хвостатых (могут утрачиваться зубы на предчелюстной и верхнечелюстной костях) зубы располагаются на верхней и нижней челюсти и на нёбной кости. У большинства бесхвостых зубы на нижней челюсти отсутствуют, а у представителей семейств настоящие жабы и узкороты, и некоторых видов из других семейств они пропадают и на верхней[42]. Считается, что зубы на нижней челюсти были утрачены ещё до выделения кроновой группы бесхвостых около 230 млн лет назад как адаптация к специфическому способу захвата пищи длинным языком. Тем не менее, у полнозубой квакши они появились вновь спустя более 200 млн лет после их утраты у предков. Таким образом, этот вид является примером нарушения закона Долло[43].

Глотка ведёт в тонкостенный пищевод. У бесхвостых он относительно короткий и широкий, а у безногих — длинный и тонкий. От глотки с одной стороны и от желудка с другой он отделён сфинктерами. Стенки пищевода имеют 6—8 продольных складок и выстланы кубическим или столбчатым ресничным эпителием с бокаловидными клетками, секретирующими слизь и пепсиноген. Из пищевода пища попадает в желудок, наиболее широкий, короткий и изогнутый у бесхвостых. Его внутренние стенки выстланы безресничным эпителием с большим количеством тонких веретеновидных клеток, секретирующих слизь. Здесь же секретируется соляная кислота, и под действием пепсина происходит расщепление белков. Из желудка пищевой комок попадает в двенадцатиперстную кишку, в которую открываются жёлчные пути и протоки поджелудочной железы. Сама поджелудочная железа лежит между двенадцатиперстной кишкой и желудком, частично окружая жёлчный проток. У хвостатых и безногих протоков поджелудочной железы два или более, а у бесхвостых лишь один. При этом у разных видов он впадает в двенадцатиперстную кишку в разных местах. Так, у жаб и квакш отверстия обоих протоков отдельные, хотя и расположены близко, у жерлянок и чесночниц отверстие общее, а у настоящих лягушек протоки сливаются ещё до впадения. Двенадцатиперстная кишка слабо отделена от тонкой, являясь её передним отделом. Стенки тонкой кишки выстланы столбчатым эпителием с бокаловидными клетками. Многоклеточные слизистые железы известны лишь у некоторых саламандр. Толстая кишка чётко отделена от тонкой лишь у некоторых бесхвостых: на границе между ними имеется клапан лоскутовидной формы. Толстая кишка переходит в клоаку, в которую открываются также половые и выделительные отверстия[44]. Печень двулопастная у бесхвостых или удлинённая у хвостатых и безногих[44]. Она имеет крупные размеры, которые у некоторых видов зависят от количества запасённого в ней гликогена и потому меняются в течение года[45].
Выделительная система
[править | править код]Личинки земноводных в качестве основного органа выделения используют головные почки, открывающиеся в целом. От них отходят вольфовые каналы, которые выводят мочу в клоаку. В ходе метаморфоза головные почки сменяются следующим поколением почек, в которых клубочки нефронов окружены боуменовыми капсулами[англ.][46]. У безногих оно представлено длинным опистонефросом[англ.] (задними почками), в формировании которого принимают участие и головные почки, а у хвостатых и бесхвостых — мезонефросом, образующимся без их участия[47]. Передняя часть почки у самцов связана с семенниками, утрачивая выделительную функцию, а вольфов канал берёт на себя также функцию проведения спермы. В связи с этим возрастает роль задней части в выделении и происходит увеличение числа нефронов в ней, что приводит к потере метамерии. Однако у некоторых представителей, например жаб-повитух и тритонов, канальцы нефронов сливаются в независимый от мюллерова протока канал, аналогичный мочеточнику амниот. У земноводных впервые появляется мочевой пузырь как карман в дне клоаки. Часто он не имеет непосредственной связи с почечными протоками. Его стенки осуществляют обратное всасывание воды, препятствуя высыханию в условиях наземной среды[46].
Земноводные, живущие во взрослом состоянии в воде, а также личинки большинства видов выделяют азот в основном в виде аммиака в больших количествах разбавленной мочи, в то время как наземные виды выделяют преимущественно мочевину, имеющую меньшую токсичность и требующую меньше воды для выведения из организма. Некоторые древесные лягушки экономят воду, выделяя до 80 % азота в виде почти нерастворимой мочевой кислоты[48].
Половая система
[править | править код]Парные половые железы подвешены на брыжейках к брюшной стороне почек[49]. К каждой гонаде прикреплены жировые тела, использующиеся в качестве источника энергии для формирования половых продуктов во время зимовки и брачного сезона. Они состоят из крупных клеток жировой ткани, окружённых тонкой оболочкой из волокнистой соединительной ткани. У самцов они как правило больше, чем у самок, и имеют более заострённые концы. Жировые тела хвостатых амфибий представляют собой длинные тяжи между гонадами и почками. У бесхвостых они находятся спереди от гонад и внешне напоминают пучки пальцевидных выростов. У безногих многочисленные листовидные жировые тела тянутся двумя рядами с каждой стороны тела от печени до клоаки. У личинок жаб между почкой и жировым телом лежит орган Биддера, который сохраняется у самцов во взрослом состоянии[50].
Половая система самца
[править | править код]
Самцы имеют парные семенники, отвечающие за сперматогенез. В брачный сезон они увеличиваются в размерах, а у некоторых бесхвостых также приобретают пигментацию. У бесхвостых семенники сферические или овальные, не разделённые на дольки. У некоторых видов они могут удлиняться, почти достигая заднего конца почки. У хвостатых почки дольчатые, причём по крайней мере у некоторых из них количество долек увеличивается с каждым успешным сезоном размножения. У безногих амфибий семенники сильно удлинённые и также состоят из долек, соединённых единым протоком. Их количество у разных видов сильно варьирует: если у крохотной червяги долька лишь одна, то у рыбозмеев их число лежит в пределах 11—20[50].
Образовавшаяся в семенниках сперма выносится по семявыносящим канальцам, лежащим в брыжейке, в ткани почки, откуда собирается вольфовым каналом и попадает в клоаку. У некоторых видов примитивных хвостатых все семявыносящие канальцы или часть из них открываются в клоаку независимо от вольфова канала[50]. На конце вольфова канала перед впадением в клоаку развивается выпячивание, называемое семенным пузырьком и служащее для хранения спермы[49].
Как правило, самец не имеет копулятивного органа[51].
Половая система самки
[править | править код]Как и семенники, женские гонады — яичники — подвешены на брыжейке к почке. Они имеют крупные веерообразно расходящиеся лопасти. Каждая лопасть окружена тонкой прозрачнойной оболочкой и состоит из гроздей яйцеклеток различной величины[52]. Когда яйцеклетки созревают, оболочка разрываются и яйцеклетки попадают в полость тела. Из целома яйцеклетки попадают в яйцеводы. Они развиваются из Мюллеровых протоков, которые в редуцированном состоянии могут встречаться и у самцов. У самцов безногих земноводных задняя часть мюллеровых протоков даже имеет железистый эпителий, который предположительно участвует в транспортировке спермы во время оплодотворения. Стенка яйцевода содержит слой гладких мышц, окружённый волокнистой соединительной тканью. Просвет яйцевода покрыт ресничным эпителием, клетки которого у живородящих видов секретируют питательные вещества для плода[50]. Яйцеводы состоят из трёх отделов: переднего, извитого и маточного. Передний отдел неизвитой и открывается воронкой, расположенной около лёгкого. Она покрыта ресничками, биение которых продвигает яйцеклетки в яйцевод. Длинный извитой отдел составляет большую часть яйцевода[52]. У яйцекладущий видов в нём имеются железы, покрывающие яйцеклетки оболочкой из пектинов[49]. Маточный отдел расширен и имеет более толстые стенки. Задним концом он открывается в клоаку[52]. В нём скапливаются икринки перед выходом наружу[49].
Нервная система
[править | править код]
В сравнении с рыбами увеличивается масса головного мозга. Так, масса головного мозга в процентах от массы тела составляет у современных хрящевых рыб 0,06—0,44 %, у костных рыб 0,02—0,94, у хвостатых земноводных 0,29—0,36, у бесхвостых 0,50—0,73 %[53].
Головной мозг земноводных относительно прост, но структурно в целом такой же, как у рептилий, птиц и млекопитающих. Их мозг удлинён, за исключением червяг, и содержит обычные двигательные и сенсорные области третрапод. Считается, что шишковидное тело, регулирующее режим сна у людей, вырабатывает гормоны, участвующие в спячке и спячке у амфибий[2].
Состоит из пяти отделов[54]:
- передний мозг относительно крупный; разделён на два полушария; имеет крупные обонятельные доли;
- промежуточный мозг хорошо развит;
- мозжечок развит слабо в связи с несложными, однообразными движениями;
- продолговатый мозг является центром дыхательной, кровеносной и пищеварительной системы;
- средний мозг относительно невелик, является центром зрения, тонуса скелетной мускулатуры.
От головного мозга отходят 10 пар черепных нервов (I—X)[54].
Спинномозговые нервы у хвостатых и бесхвостых образуют хорошо выраженные плечевое и поясничное сплетения. Хорошо развита симпатическая нервная система, представленная в основном двумя симпатическими нервными стволами, расположенными по бокам позвоночного столба[54]. Спинной мозг имеет парные корешки спинномозговых нервов: задний — чувствительный, и передний — двигательный. Их количество варьируется у различных групп амфибий, например у бесхвостых их обычно 10-11 пар. У взрослых представителей двух других отрядов современных амфибий их количество больше, как и у личинок, теряющих хвосты и развивающих конечности[54]. Средний мозг у представителей семейств настоящие лягушки и жабы занимает 21 % головного мозга, у представителей рода Aмбистомы только 7 %. Он состоит из двух основных частей. Верхнее двухолмие лежит дорсально, а покрышка среднего мозга вентрально. В первом есть области, отвечающие за зрение и слух, которые служат для интеграции поступающих сигналов. Они существенно отличаются у отрядов амфибий. У лягушек эти участки довольно хорошо развиты, в отличие от представителей безногих и хвостатых отрядов. В этом сегменте расположены некоторые ядра черепно-мозговых нервов. Средний мозг иногда считают самой крупной и «высокой» частью мозга земноводных, что явно ошибочно[54]. Обонятельная доля участвует в обработке запаха (большая часть переднего мозга занимается обработкой обонятельных стимулов). Головная кора отвечает за интеллект и контролирует поведение. Несмотря на схожесть с органами млекопитающих, эти органы выполняют совершенно другие функции. Основным отличием является также отсутствие новой коры; её функции, по-видимому, выполняет дорсальная часть плаща мозга. В пределах промежуточного мозга можно выделить эпиталамус, таламус, обрабатывающий информацию от органов чувств, отличных от обонятельных, и гипоталамус, который, в свою очередь, соединяется с гипофизом и контролирует секрецию[54]. Спереди от промежуточного мозга лежит мозжечок. Мозжечок амфибий меньше, чем у любых других позвоночных, за исключением нижнечелюстных, и отвечает за координацию мышц[54].
Органы чувств
[править | править код]Органы зрения
[править | править код]_(6941089388).jpg)
Глазные яблоки современных земноводных имеют почти сферическую форму. Спереди они покрыты прозрачной мембраной, роговицей[55], которая имеет более выпуклую форму по сравнению с плоской роговицей рыб[56]. Остальная их часть покрыта плотной соединительнотканной оболочкой, склерой. У бесхвостых и большинства личинок хвостатых она поддерживается хрящом в форме кольца или чаши, особенно развитым у скрытожаберников, некоторых наземных хвостатых и узкорота Stereocyclops incrassatus. Снаружи глаз прикрывается веками, которые развиваются в ходе метаморфоза у всех земноводных, кроме безногих, у которых глаза погружаются под полупрозрачный слой кожи или кость, пиповых и облигатно неотенических саламандр. Верхнее веко представляет собой малоподвижную кожную складку. Нижнее веко более подвижно и в сложенном состоянии образует N-образную складку. Верхняя её часть представляет собой тонкую полупрозрачную мигательной перепонки[55].
Смачивание глаза происходит за счёт выделений различных экзокринных желёз, попадающих на конъюнктиву. У некоторых хвостатых эти железы дифференцируются на гардеровы железы, расположенные спереди и смачивающие мигательную перепонку, и слёзные железы, расположенные по бокам глаза. У бесхвостых и безногих имеются лишь гардеровы железы, причём у последних они смачивают не глаза, а чувствительные щупальца[55].
Хрусталик крупный и уплощённый[55], имеет форму двояковыпуклой линзы[56], но его уплощение происходит в ходе метаморфоза, потому у облигатно неотенических саламандр он круглый. У червяг хрусталик мутный. Его аккомодация осуществляется цилиарным телом, наиболее развитым у бесхвостых и отсутствующим у безногих. Хрусталик окружён пигментированной радужной оболочкой. У хвостатых и бесхвостых она состоит из двух сетчатковых слоёв и одного слоя стромы. В каждом из сетчатковых слоёв присутствуют пигментные клетки — иридофоры и меланофоры, а у некоторых бесхвостых также ксантофоры. У безногих в радужке строма отсутствует, а пигментные клетки имеются только в одном из сетчатковых слоёв. Радужка обрамляет зрачок и регулирует количество проходящего через него света за счёт работы дилататора и сфинктера зрачка. Зрачок хвостатых земноводных круглый, а у бесхвостых он может быть также треугольным, вертикальным или горизонтальным[55].

Сетчатка состоит из пяти слоёв: пигментного эпителия, наружного ядерного, наружного сетчатого, внутреннего ядерного и внутреннего сетчатого. Зрительные рецепторы земноводных представлены четырьмя видами: зелёными и красными палочками и простые и двойные колбочками[55]. Амфибии являются единственным классом позвоночных, имеющим два типа палочек, при этом зелёные палочки у других животных не встречаются[57].
У ископаемых беспанцирных земноводных имелся хорошо различимый теменной глаз[58]. Он отсутствует у хвостатых и безногих, но сохранился у некоторых бесхвостых под кожей между глазами[59][60].
Органы обоняния и вкуса
[править | править код]Органы обоняния представлены парными носовыми мешками, расположенными в передней части черепа. Каждый мешок имеет по две ноздри — наружную, открывающуюся в окружающую среду, и внутреннюю — хоану — ведущую в ротоглоточную полость. Появление внутренних ноздрей позволило включить обонятельные полости в состав дыхательных путей[61]. Носовой мешок состоит из основной носовой полости и добавочных полостей, расположенных вентрально и латерально. Одной из них является вомероназальный орган. От переднего края глаза к передней части основной носовой камеры идёт носослёзный проток[57]. У хвостатых и особенно у бесхвостых носовые мешки поддерживаются хрящём, в то время как у безногих, у которых хорошо развиты крыша черепа и нёбный комплекс, носовые хрящи развиты слабо[62].

Стенки мешков выстланы эпителием двух видов: дыхательным и обонятельным, а также железами, выделяющими слизь. Дыхательный эпителий находится на медиальной и латеральной стенках основной носовой полости. Биение ресничек клеток эпителия, поддерживает ток слизи по нему. Обонятельный эпителий залегает в трёх зонах: на медиальной стенке, крыше и переднем конце основной носовой полости; на приподнятой части дна основной носовой полости и в вомероназальном органе, расположенном в отдельной полости. Первые две зоны иннервируются дорсальной и вентральной ветвями обонятельного нерва соответственно, а третья зона — собственным вомероназальным нервом[62].
У хвостатых земноводных носовые мешки устроены наиболее просто. Основная носовая полость у них крупная и разделена на две половины продольной складкой. У водных хвостатых происходит упрощение носовых мешков, и отдельные их части редуцируются. Так, у сиренов отсутствует носослёзный проток, у протеев и скрытожаберников — вомероназальный орган, а у амфиум — и то, и другое[62]. У бесхвостых носовые мешки усложняются: основные носовые полости дифференцируются на три камеры, а вомероназальный орган увеличивается. У безногих носовые мешки по строению приближены к мешкам хвостатых, однако у них имеются чувствительные щупальца, которые могут выдвигаться наружу. Через носослёзный проток они передают пахучие частицы вомероназальному органу[57].
Во рту и глотке[63], а также в коже расположены вкусовые луковицы. Предполагается, что они способны распознавать только горький и солёный вкусы[64].
Органы осязания
[править | править код]
Исходными для земноводных органами осязания являются органы боковой линии. Они присутствуют у водных личинок и у взрослых земноводных, живущих в воде, таких как скрытожаберники, амфиумы, сирены, пиповые[65] и водные червяги[66]. У земноводных встречаются два типа органов боковой линии: невромасты, имеющие механорецепторную функцию, и ампулы Лоренцини, отвечающие за электрорецепцию[65].
Невромасты могут располагаться поодиночке (например у сиренов) или небольшими рядами вдоль латеральной и дорсолатеральной поверхностей тела. Особенно заметны они на голове, где могут формировать отчётливый рисунок. Они состоят из группы клеток, имеющей грушевидную форму и погружённой в эпидермис. У личинок невромасты выступают над поверхностью кожи, в то время как у взрослых земноводных они обычно расположены в небольших углублениях. Каждый невромаст состоит из трёх типов клеток: клеток мантии, окружающих остальные клетки невромаста по периферии, поддерживающих клеток, залегающих непосредственно на базальной мембране, и волосковых клеток, лежащих на поддерживающих клетках в апикальной части невромаста. Каждая волосковая клетка имеет по одному длинному волоску и множеству более коротких волосков, длина которых уменьшается при удалении от длинного. Волоски невромаста покрыты тонкой лентовидной купулой. Каждый невромаст иннервируется одним эфферентным и двумя афферентными нервными волокнами. Большая часть невромастов головы иннервируется передним нервом боковой линии (отходит от лицевого нерва), а остальные — задним нервом боковой линии (отходит от блуждающего нерва)[65].
Ампулы Лоренцини встречаются у личинок безногих и водных хвостатых земноводных. Они расположены на голове, где расположены в ряды, параллельные рядам невромастов. По строению они напоминают невромасты, но отличаются тем, что погружены в эпидермис своей апикальной частью, не имеют купулы и вместо волосков на чувствительных клетках расположены микроворсинки. Каждая ампула Лоренцини иннервируется лишь одним афферентным нервным волокном[65].
Помимо органов боковой линии в коже земноводных присутствуют нервные окончания, воспринимающие холод и тепло (в эпидермисе), а также боль и давление (в дерме)[65].
Органы слуха
[править | править код]
В процессе эволюции земноводные приобрели орган слуха, способный воспринимать как вибрации субстрата, так и колебания воздуха[67]. У большинства бесхвостых орган слуха состоит из двух отделов: внутреннего и среднего уха. Однако у некоторых представителей, например у Odorrana tormota, развивается также наружное ухо в виде небольшой полости перед барабанной перепонкой[68]. У хвостатых и безногих имеется лишь внутреннее ухо[67].
Среднее ухо бесхвостых представляет собой воздушную полость между барабанной перепонкой и мембраной овального окна, ведущего во внутреннее ухо. В нём имеется слуховая косточка, стремечко. Своей дистальной хрящевой частью, экстраколумеллой, стремечко упирается в центр барабанной перепонки, передавая колебания от неё к овальному окну[68]. С овальным окном связан также костный (у некоторых хвостатых) или хрящевой (у остальных хвостатых и бесхвостых) элемент — оперкулум[69]. С помощью специальной мышцы он связан с надлопаточным хрящом, через переднюю конечность принимая колебания от субстрата[70]. Полость среднего уха соединена с глоткой евстахиевой трубой, что позволяет выравнивать давление по обе стороны от барабанной перепонки. У хвостатых и безногих среднее ухо и евстахиева труба отсутствуют, и стремечко своим наружным концом соединяется с щёчной областью черепа или полностью редуцируется[71].
Внутреннее ухо лежит в ушной капсуле, поддерживаемое рыхлой соединительной тканью. Оно состоит из двух систем мембран, заполненных жидкостью: перилимфатической и эндолимфатической. Перилимфатическая система передаёт колебания от овального окна на эндолимфатическую систему, которая состоит из двух основных мешочков: дорсального овального (лат. utriculus) и вентрального круглого (лат. sacculus)[67][71]. От овального мешочка отходят три полукружных канала В свою очередь, от круглого мешочка отходят три структуры: выступ (лат. lagena) от постеровентральной стенки, амфибийный сосочек (лат. papilla amphibiorum) от дорсальной и базилярный сосочек (лат. papilla basilaris) от вентральной[67][71]. В этих структурах расположены участки чувствительного эпителия, иннервируемые задней ветвью слухового нерва и воспринимающие различные колебания. Клетки чувствительного эпителия внутреннего уха снабжены волосками, как клетки невромастов боковой линии. В полукружных каналах и овальноми мешочке (имеется только у безногих) они покрыты купулой[67] и воспринимают угловое и линейное ускорение головы соответственно[68]. Другие участки чувствительного эпителия расположены в овальном и круглом мешочках, лагене и сосочках. Они отличаются наличием отолитов в купуле. Амфибийный сосочек встречается только у земноводных[67]. Его чувствительный эпителий воспринимает звуковые волны низкой частоты, в то время как эпителий базилярного сосочка — высокой[68].
Примечания
[править | править код]- ↑ 1 2 Дзержинский и др., 2013, с. 197—198.
- ↑ 1 2 Stebbins, 1995.
- ↑ 1 2 Vitt & Caldwell, 2014, p. 48.
- ↑ Ромер, Парсонс, 1992, с. 166.
- ↑ 1 2 3 4 5 6 7 Duellman, Trueb, 1994, p. 368—376.
- ↑ 1 2 3 Fox H. Chapter 5. Epidermis // Biology of the Integument : [англ.] / Edited by Bereiter-Hahn, A. G. Matoltsy, K Sylvia Richards. — Springer-Verlag Berlin Heidelberg, 1986. — Vol. 2. Vertebrates. — P. 97—105. — 855 p. — ISBN 978-3-662-00989-5.
- ↑ Edwin Nicholas Arnold. A field guide to the reptiles and amphibians of Britain and Europe. — London: Collins, 1978. — 272 pages, 40 pages of plates с. — ISBN 0-00-219318-3, 978-0-00-219318-4.
- ↑ Гуртовой и др., 1978, с. 11.
- ↑ 1 2 Fox H. Chapter 6. Dermis // Biology of the Integument : [англ.] / Edited by Bereiter-Hahn, A. G. Matoltsy, K Sylvia Richards. — Springer-Verlag Berlin Heidelberg, 1986. — Vol. 2. Vertebrates. — P. 111—114. — 855 p. — ISBN 978-3-662-00989-5.
- ↑ R. I. C. Spearman. The integument: a textbook of skin biology,. — London,: Cambridge University Press, 1973. — vii, 208, [8] pages с. — ISBN 0-521-20048-2, 978-0-521-20048-6.
- ↑ Richard Ian Campbell Spearman. The Integument. — CUP Archive, 1973-06-28. — 232 с. — ISBN 978-0-521-20048-6.
- ↑ 1 2 Robert L. Dorit, 1991, p. 843—878.
- ↑ 1 2 3 4 5 Дзержинский и др., 2013, с. 200—202.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Наумов, Карташёв, 1979, с. 282—288.
- ↑ Константинов, Шаталова, 2005, с. 66—67.
- ↑ 1 2 3 Vitt & Caldwell, 2014, p. 59—60.
- ↑ 1 2 3 4 5 6 7 Vitt & Caldwell, 2014, p. 62—64.
- ↑ Константинов, Шаталова, 2005, с. 95—96.
- ↑ Гуртовой и др., 1978, с. 77.
- ↑ Константинов, Шаталова, 2005, с. 110—112.
- ↑ Гуртовой и др., 1978, с. 126—141.
- ↑ 1 2 3 4 5 6 Stebbins, 1995, p. 17—25.
- ↑ 1 2 3 4 5 6 7 Наумов, Карташёв, 1979, с. 291—293.
- ↑ 1 2 Vitt & Caldwell, 2014, p. 76—77.
- ↑ 1 2 Jean-Marie Exbrayat. Reproductive Biology and Phylogeny of Gymnophiona (Caecilians) : [англ.] / Series Editor: Barrie G. M. Jamieson. — The Science Publishers, 2006. — Vol. 5. — P. 96—97. — 395 p. — (Reproductive Biology and Phylogeny Series). — ISBN 1-57808-312-5 978-1-57808-312-1.
- ↑ Дзержинский и др., 2013, с. 207—208.
- ↑ Ульяхин А. Лягушка без легких. Элементы.ру (23 мая 2018). Дата обращения: 27 октября 2022. Архивировано 27 октября 2022 года.
- ↑ 1 2 3 4 5 Наумов, Карташёв, 1979, с. 293—297.
- ↑ Константинов, Шаталова, 2005, с. 172—176.
- ↑ 1 2 Гуртовой и др., 1978, с. 28—40.
- ↑ 1 2 3 Duellman, Trueb, 1994, p. 398—399.
- ↑ 1 2 Дзержинский и др., 2013, с. 211—212.
- ↑ Ромер, Парсонс, 1992, с. 181—184.
- ↑ 1 2 3 Ромер, Парсонс, 1992, с. 143—153.
- ↑ Villolobos M., León P., Sessions S. K., Kezer L. Enucleated Erythrocytes in Plethodontid Salamanders : [англ.] : [арх. 5 марта 2023] // Herpetologica. — 1988. — Vol. 44, no. 2. — P. 243—250.
- ↑ 1 2 3 Ромер, Парсонс, 1992, с. 15—21.
- ↑ Константинов, Шаталова, 2005, с. 118.
- ↑ 1 2 Stebbins, 1995, p. 57—61.
- ↑ Наумов, Карташёв, 1979, с. 289.
- ↑ Константинов, Шаталова, 2005, с. 123.
- ↑ Davit-Béal T., Chisaka H., Delgado S., & Sire J.-Y. Amphibian teeth: current knowledge, unanswered questions, and some directions for future research : [англ.] // Biological Reviews. — 2007. — Vol. 82. — P. 49–81. — doi:10.1111/j.1469-185X.2006.00003.x.
- ↑ Paluh D. J., Riddell K., Early C. M., Hantak M. M., Jongsma G. F. M., Keeffe R. M., Silva F. M., Nielsen S. V., Vallejo-Pareja M. C., Stanley E. L., Blackburn D. C. Rampant tooth loss across 200 million years of frog evolution : [арх. 30 ноября 2022] // eLife. — 2021. — Vol. 10. — P. e66926. — doi:10.7554/eLife.66926.
- ↑ Wiens, J. J. Re-Evolution of Lost Mandibular Teeth in Frogs After More Than 200 Million Years, and Re-Evaluating Dollo's Law // Evolution. — 2011. — Vol. 65. — P. 1283—1296. — doi:10.1111/j.1558-5646.2011.01221.x. Архивировано 9 октября 2022 года.
- ↑ 1 2 Duellman, Trueb, 1994, p. 408—411.
- ↑ Stebbins, 1995, p. 66.
- ↑ 1 2 Ромер, Парсонс, 1992, с. 101—115.
- ↑ Duellman, Trueb, 1994, p. 405—407.
- ↑ Robert L. Dorit, 1991, с. 849.
- ↑ 1 2 3 4 Дзержинский и др., 2013, с. 213—214.
- ↑ 1 2 3 4 Duellman, Trueb, 1994, p. 407—408.
- ↑ Dev Raj Khanna, P. R. Yadav. Biology of Amphibia. — Discovery Publishing House, 2005. — С. 7. — 412 с. — ISBN 978-81-7141-932-6.
- ↑ 1 2 3 Гуртовой и др., 1978, с. 27—28.
- ↑ Никитенко, 1969. Головной мозг земноводных. Биология и медицина. Дата обращения: 13 марта 2012. Архивировано 16 декабря 2013 года.
- ↑ 1 2 3 4 5 6 7 Martin E. Feder, Warren W. Burggren. Environmental Physiology of the Amphibians. — University of Chicago Press, 1992-10-15. — 660 с. — ISBN 978-0-226-23944-6.
- ↑ 1 2 3 4 5 6 7 Duellman, Trueb, 1994, p. 379—382.
- ↑ 1 2 Наумов, Карташёв, 1979, с. 310.
- ↑ 1 2 3 Vitt & Caldwell, 2014, p. 70.
- ↑ Kardong, 2018, p. 691.
- ↑ Duellman, Trueb, 1994, p. 383—384.
- ↑ Vitt & Caldwell, 2014, p. 80.
- ↑ Дзержинский и др., 2013, с. 205.
- ↑ 1 2 3 Duellman, Trueb, 1994, p. 384—386.
- ↑ Kardong, 2018, p. 679.
- ↑ Ромер, Парсонс, 1992, с. 202—203.
- ↑ 1 2 3 4 5 Duellman, Trueb, 1994, p. 378—379.
- ↑ Vitt & Caldwell, 2014, p. 68.
- ↑ 1 2 3 4 5 6 Duellman, Trueb, 1994, p. 387—390.
- ↑ 1 2 3 4 Schoffelen R. L. M., Segenhout J. M. & van Dijk P. Mechanics of the exceptional anuran ear : [англ.] // Journal of Comparative Physiology A. — 2008. — Vol. 194. — P. 417–428. — doi:10.1007/s00359-008-0327-1.
- ↑ Lewis E. R. & Narins P. M. The Acoustic Periphery of Amphibians: Anatomy and Physiology // Comparative Hearing: Fish and Amphibians : [англ.]. — New York : Springer, 1999. — Vol. 11. — P. 101–154. — 438 p. — (Springer Handbook of Auditory Research). — ISBN 978-1-4612-0533-3 (online). — doi:10.1007/978-1-4612-0533-3_4.
- ↑ Kardong, 2018, p. 704.
- ↑ 1 2 3 Ромер, Парсонс, 1992, с. 244—246.
Литература
[править | править код]- Гуртовой Н. Н., Матвеев Б. С., Дзержинский Ф. Я. Практическая зоотомия позвоночных. Земноводные, пресмыкающиеся. — М.: Высшая школа, 1978. — 407 с.
- Дзержинский Ф. Я., Васильев Б. Д., Малахов В. В. Зоология позвоночных. — М.: Академия, 2013. — 464 с. — ISBN 978-5-7965-7971-4.
- Наумов Н. П., Карташёв Н. Н. Зоология позвоночных. — М.: Высшая школа, 1979. — Т. 1. Низшие хордовые, бесчелюстные, рыбы, земноводные. — 333 с.
- Ромер А., Парсонс Т. Анатомия позвоночных / пер. с англ. А. Н. Кузнецова и В. Б. Никитина. — М.: Мир, 1992. — Т. 2. — 406 с. — ISBN 5-03-000292-8.
- Константинов В. М., Шаталова С. П. Сравнительная анатомия позвоночных животных. — М.: Академия, 2005. — 304 с. — ISBN 5-7695-1770-0.
- Duellman W. E., Trueb L. Biology of amphibians (англ.). — Baltimore: Johns Hopkins University Press, 1994. — 670 p.
- Grzimek, B. Grzimek’s Animal Life Encyclopedia (англ.). — 2nd edition. — Farmington Hills: Gale Group[англ.], 2003. — Vol. 6, Amphibians. — P. 3—590. — ISBN 0-7876-5362-4. (недоступная ссылка)
- Kardong K. V. Vertebrates: Comparative Anatomy, Function, Evolution. — 8. — New York: McGraw-Hill Education, 2018. — 796 p. — ISBN 978-1-259-70091-0. — ISBN 978-1-260-39856-4.
- Robert L. Dorit. Zoology. — Philadelphia: Saunders College Pub, 1991. — 1126 с.
- Robert C. Stebbins. A natural history of amphibians. — Princeton, N.J.: Princeton University Press, 1995. — xvi, 316 pages с. — ISBN 0-691-03281-5, 978-0-691-03281-8, 0-691-10251-1, 978-0-691-10251-1.
- Vitt L. J., Janalee P. Caldwell. Herpetology : an introductory biology of amphibians and reptiles. — 4th edition. — Amsterdam, 2014. — xiv, 757 pages с. — ISBN 978-0-12-386919-7, 0-12-386919-6.
Дополнительная литература
[править | править код]- Жизнь животных. В 7 т. / гл. ред. В. Е. Соколов. — 2‑е изд., перераб. — М. : Просвещение, 1985. — Т. 5 : Земноводные. Пресмыкающиеся / под ред. А. Г. Банникова. — 399 с. : ил.
- Robert F. Inger, Robert B. Stuebing: A Fieldguide to the Frogs of Borneo. Natural History Publications (Borneo), Kota Kinabalu 2005, ISBN 983-812-085-5.
- Green DM, Sessions SK (1991) Amphibian cytogenetics and evolution. Academic Press, San Diego
- King M. (1990) Animal cytogenetics: Amphibia. Gebruder Borntraeger, Berlin
- Beltz, Ellin. Frogs: Inside their Remarkable World (англ.). — Firefly Books, 2005. — ISBN 1-55297-869-9.
- Cogger, H.G.; R.G. Zweifel, and D. Kirschner. Encyclopedia of Reptiles & Amphibians Second Edition (англ.). — Fog City Press, 2004. — ISBN 1-877019-69-0.
- Tyler, M. J. Australian Frogs A Natural History (англ.). — Reed Books[англ.], 1994. — ISBN 0-7301-0468-0.
- Blackburn D. C., Wake D. B. Class Amphibia Gray, 1825 // Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness / Zhi-Qiang Zhang (editor). — Auckland: Magnolia Press, 2011. — P. 39—55. — 237 p. — (Zootaxa). — ISBN 1175-5334.
- Grzimek B. Grzimek’s Animal Life Encyclopedia[англ.] / William E. Duellman, Advisory Editor. — 2nd edition. — Farmington Hills[англ.]: Gale Group[англ.], 2003. — P. 3—321. — ISBN 0-7876-5362-4.