Малоазиатский тритон (Bglkg[ngmvtnw mjnmku)
| Малоазиатский тритон | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||
| Научная классификация | ||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Класс: Подкласс: Инфракласс: Отряд: Подотряд: Семейство: Подсемейство: Вид: Малоазиатский тритон |
||||||||||
| Международное научное название | ||||||||||
|
Ommatotriton ophryticus (Berthold, 1846) |
||||||||||
| Синонимы | ||||||||||
|
||||||||||
| Охранный статус | ||||||||||
| ||||||||||

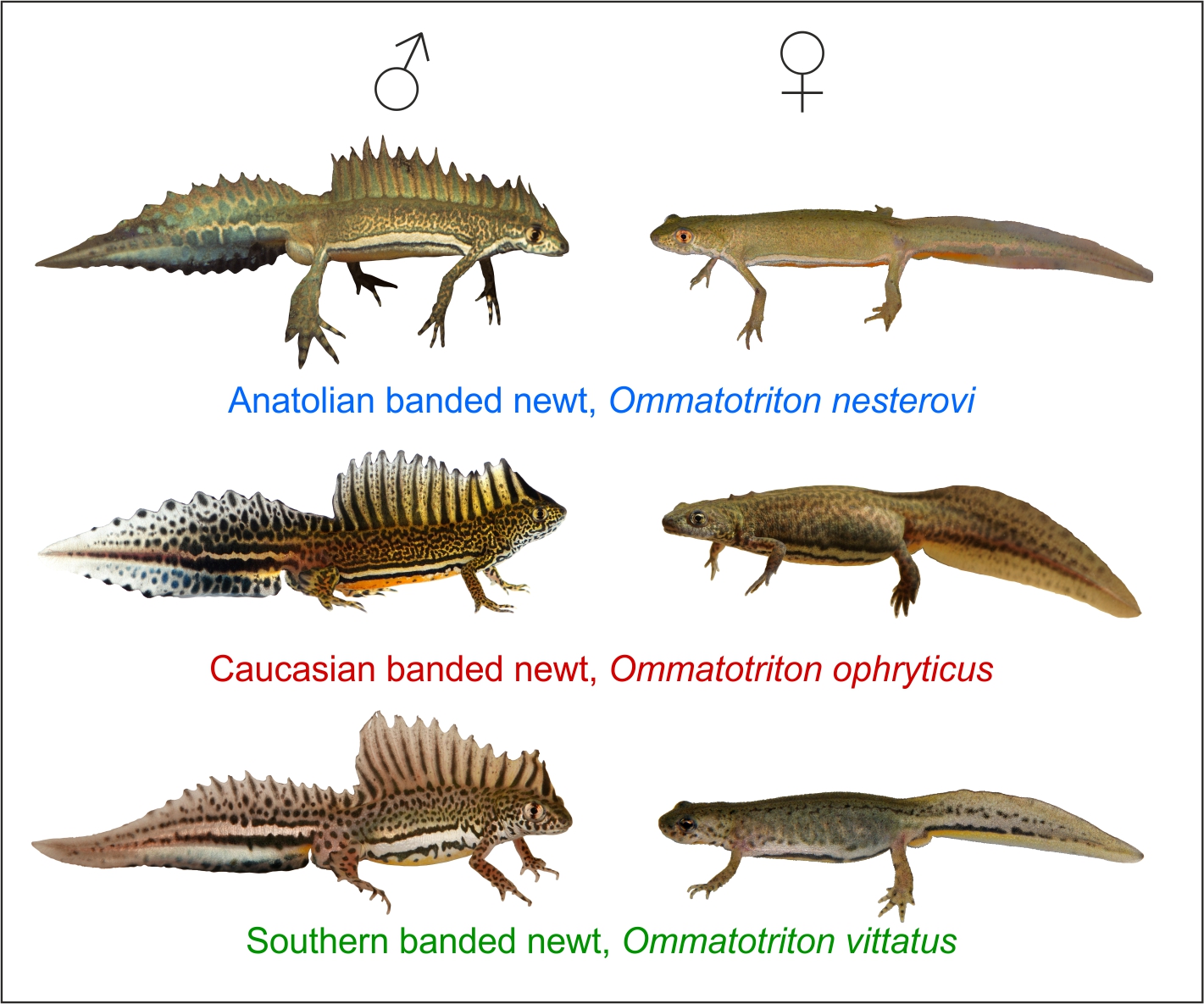
Малоазиа́тский трито́н[2][3] (лат. Ommatotriton ophryticus) — вид хвостатых земноводных из подсемейства Pleurodelinae семейства настоящих саламандр. Эндемик Восточного Причерноморья, обитает в предгорных и горных лесах и на альпийских лугах Западного Кавказа на юг до северо-востока Малой Азии в основном на высоте от 1200 до 1500 м. Относительно крупный тритон длиной до 17 см и с кирпично-красноватой окраской.
На спине у самцов высокий гребень, в брачный период, когда тритоны живут в водоёмах, увеличивающийся ещё больше, до 3 см в высоту. Во время размножения самцы становятся яркоокрашенными, золотисто-жёлтыми или бронзово-оливковыми с многочисленными мелкими тёмными пятнышками на спине и боках, жёлто-оранжевым брюхом и яркой серебристо-белой полосой, окаймлённой сверху и снизу тёмными линиями. Размножение в зависимости от высоты обитания происходит с марта в предгорьях до июля на высокогорье. Для этого тритоны спускаются в водоёмы, стоячие или слабопроточные, но обязательно богатые водной растительностью или с устланным опавшими листьями дном.
Особенно тритоны любят крупные горные озёра с чистой и бедной питательными веществами водой, в которых не водится рыба и где икре и личинкам можно безопасно развиваться. Икру откладывают на подводные растения или листья, по одной икринке или короткими цепочками. Икринки сравнительно крупные, до 2—4 мм в диаметре. Через 2—4 недели из них появляются личинки, которые держатся среди растительности и питаются мелкими водными беспозвоночными, в основном ракообразными. Взрослые тритоны в водоёмах также питаются водными, а на суше наземными беспозвоночными, употребляя также икру и головастиков других амфибий.
Редкий вид с численностью, сокращающейся из-за осушения и загрязнения водоёмов, браконьерского отлова и хищничества завезённого человеком енота-полоскуна. Занесён в Красные книги всех стран, на территории которых встречается. Охраняется в нескольких заповедниках и национальных парках. Однако для стабилизации популяции вида необходимы более строгая охрана вида и мест его обитания, более широкое внедрение технологии содержания и разведения его в неволе для реинтродукции в природу.
Описание
[править | править код]

Относительно крупный тритон, общая длина тела с хвостом может превышать 17 см[4][5][6], но обычно лежит в пределах 10,6—15,8 см, без хвоста — 5,1—8,8 см. Хвост по длине равен длине туловища с головой либо немного больше или меньше[2]. Масса тела до примерно 14 г. С увеличением высоты обитания длина и масса тела тритонов уменьшаются[7]. Во многих популяциях самцы крупнее самок[2]. Наиболее крупные особи встречаются в среднегорье южного склона Главного Кавказского хребта[4]. Особи с Северо-Западного Кавказа в целом крупнее, чем таковые из Восточной Грузии; тритоны с высокогорий крупнее особей, обитающих на более низких высотах в пределах одного и того же региона[2]. Конечности и пальцы относительно длинные. На передних конечностях по четыре пальца, на задних по пять. Третий палец передней конечности иногда длиннее предплечья, а четвёртый палец задней конечности — бедра[8]. Кожа тритонов во время водного и наземного периодов жизни отличается по своему внешнему виду: в брачный сезон, когда эти животные держатся в воде, она гладкая, глянцевая, слизистая и хорошо смачивается водой; в период обитания на суше — бархатистая, напоминающая замшу, слабо зернистая, плотная, полностью лишённая слизи и плохо смачивается водой[4][5][9]. Паротиды не выступающие. Костальные (рёберные) борозды развиты слабо[8]. Окраска спины и боков во время наземной фазы жизни кирпично- или оранжево-красноватая, оранжевая или оранжево-бурая с двумя округлыми жёлтыми пятнами на верхней стороне головы[5][4]. В период размножения, когда тритоны обитают в воде, самцы становятся яркоокрашенными: спина и бока от золотисто-жёлтого до бронзово-оливкового и оливково-бурого цвета с многочисленными мелкими тёмными пятнышками, на боках внизу между передними и задними конечностями проходит узкая продольная серебристо-белая полоса, окаймлённая сверху и снизу тёмными линиями. Горло пятнистое. Брюхо от белёсо-жёлтого до яично-жёлтого и оранжевого цвета, однотонное. Вдоль середины спины на всём её протяжении у самцов расположен сильно зазубренный по верхнему краю гребень, прерывающийся у основания хвоста. В период размножения он увеличивается, достигая в высоту иногда более 3 см. Увеличивается он значительнее, чем у других видов тритонов. Гребень желтоватого или коричневатого цвета с тёмно-зелёными вертикальными полосами, которых может быть до 14, идущими от спины к вершинам «зубцов» гребня[8][10][5][4]. Хвост уплощён с боков и по форме напоминает лист[6]. Во время брачного периода у самцов верхняя сторона хвоста такого же цвета, что и верх туловища, с чёрно-белым сетчатым рисунком, нижняя — ярко-синяя или перламутрово-синяя, в передней части часто с конусообразными тёмно-синими продольными полосками[10], снизу и с боков с голубыми и/или зеленоватыми пятнами[2]. Вдоль середины хвоста проходят два продольных ряда чёрных пятен, которые иногда сливаются в продольные полосы. Самки окрашены менее ярко, в их окраске преобладают буро-оливковые тона с ярко-жёлтым или оранжевым брюхом и слабо выраженной боковой серебристой полосой[10], тёмные пятна у них почти полностью отсутствуют, спинного гребня нет[6]. Молодые тритоны, покидающие водоём после метаморфоза (сеголетки), окрашены в кофейно-жёлтые, бурые и кирпично-красные тона с двумя охристыми затылочными пятнами[4] и слабовыраженным рисунком на теле[6]. По мере наступления полового созревания окраска меняется и становится характерной для каждого пола[4].
{kind=link}
В пределах ареала с малоазиатским тритоном в одних и тех же биотопах обитают три вида хвостатых амфибий. От кавказского обыкновенного тритона, или тритона Ланца, он отличается окраской и формой тела (в первую очередь высоким и прерывающимся у хвоста спинным гребнем у самцов), от тритона Карелина яркой окраской и отсутствием пятен на брюхе, от кавказской саламандры формой и окраской тела и хвоста[5]. Основным морфологическим признаком, позволяющим отличить малоазиатского тритона от двух других видов рода Ommatotriton, является продольная белая полоса внизу боков тела. У ближневосточного тритона эта боковая белая полоса заметно шире, чем у малоазиатского. У вида Ommatotriton nesterovi она тянется не только между передними и задними конечностями, но и от передних конечностей до глаза, не прерываясь пятнами или крапинами. Однако узкая продольная светлая полоса на хвосте у Ommatotriton nesterovi прерывается крупными пятнами (у самцов они с голубоватым оттенком), в то время как у Ommatotriton ophryticus она непрерывная[11].
Кариотип малоазиатского тритона представлен 12 парами хромосом, диплоидное число хромосом 2n = 24, основное число хромосом NF = 48. Размер генома 66,8—68,8 пикограмма[2]. В позвоночнике 13 предкрестцовых (туловищных) позвонков[12].
Ареал
[править | править код]Восточнопонтийский эндемик. Малоазиатский тритон распространён на крайнем юго-востоке Европы в Восточном Причерноморье: на юге Краснодарского края, в Адыгее, Карачаево-Черкесии, на Западном Кавказе и на северо-востоке Малой Азии. На Кавказе встречается на северо-западном и юго-западном макросклонах Главного Кавказского хребта. На севере ареал вида ограничен границами лесного пояса Кавказских гор[3]. Северо-восточная граница ареала проходит в районе восточного берега реки Кубань в Предкавказье. На восток распространён до Южной Осетии и Лагодехи[13], с востока ареал ограничен западными склонами горы Эльбрус[3]. Возможно, встречается на крайнем западе Азербайджана. К югу от Главного Кавказского хребта ареал вида охватывает Грузию от черноморского побережья до городов Телави и Тбилиси, крайний север Армении в пределах севера Лорийской области[14] и восточную часть причерноморского побережья Турции[13][15]. Многие популяции на периферии ареала являются географически изолированными. В ареал вида глубоко врезается сухая, а потому недоступная для тритонов, долина реки Кура[13]. Известны случаи находок малоазиатского тритона в некоторых местах в центральной части Краснодарского края и в Ставропольском крае, однако они, скорее всего, связаны с интродукцией туда этого вида, и эти регионы не входят в его естественный ареал[16][3]. Вид интродуцирован также на северо-востоке Испании в Каталонии, где теперь обитает гибридная популяция видов Ommatotriton ophryticus и Ommatotriton nesterovi[15][1]. Малоазиатский тритон предпочитает мягкий климат без резких перепадов температуры. На его распространение также оказывает значительное влияние количество осадков в наиболее сухом месяце[15].
Места обитания
[править | править код]
Горно-лесной и горно-луговой вид, связанный с влажными горными лесами и лугами и лишь изредка проникающий на предгорные равнины. Обитает преимущественно в горных хвойных, смешанных и лиственных лесах, в которых преобладают берёза, дуб, граб, ольха, каштан, бук, рододендрон и т. д., в основном на высотах от 1200 до 1500 м. В целом же встречается от приморских низменностей (например, в районах Адлера и Пицунды, где высота почти 0 м над уровнем моря) с влажным субтропическим климатом до нижнеальпийского пояса, до высоты 2743 м. На востоке ареала, где климат более сухой, обитает на высотах 600—2050 м. На Западном Кавказе обитает в лесах всех типов: в широколиственных субтропических лесах с вечнозелёным подлеском, каштанниках, букняках мертвопокровных, прирусловых и ольхово-ивовых лесах, тисо-самшитовых рощах (самшитниках), ясеневых букняках. Встречается также в кустарниковых зарослях, растущих на месте сведённых лесов, на обезлесенных склонах гор, в агроценозах, на высокогорье — у верхней границы леса, в берёзовых рощах и среди рододендроновых формаций, на субальпийских и альпийских лугах[13][3][15][10][5]. В Восточном Закавказье встречается в колхидских рефугиумах[4]. Малоазиатский тритон предпочитает хорошо увлажнённые биотопы с большим количеством слабопроточных и стоячих водоёмов[15]. Иногда встречается в карстовых пещерах, куда попадает с потоками воды. Самая «глубокая» находка была сделана в Назаровской пещере (хребет Алек на северо-западе Кавказа), где тритона обнаружили на расстоянии 400 м от входа в пещеру[17][13][3].
Всего в пределах естественного ареала известно около 317 локалитетов обитания малоазиатского тритона: 151 в предгорных и горных районах Предкавказья и северо-запада Кавказа, 111 в Западном Закавказье и 55 на северо-востоке Малой Азии[15].
В кавказской части ареала в одних биотопах с малоазиатским тритоном обитают тритон Ланца и тритон Карелина (синтопичные виды)[18], все три вида размножаются в одних и тех же водоёмах, их популяционные ареалы зачастую практически совпадают[19]. Однако малоазиатский тритон более устойчив к низкой влажности, чем тритон Ланца, и менее чувствителен к размеру и глубине водоёмов, чем тритон Карелина[20].
Популяция
[править | править код]Образует полуизолированные или изолированные популяции, использующие для размножения отдельные водоёмы или их группы в горах[13]. Ранее в водоёмах размножения отмечалось 4—6 особей на 1 м². В окрестностях Гузерипля по реке Жёлобной отмечали крупные скопления в небольших водоёмах в буко-пихтарниках, до 100 особей на 25 м². В отдельных крупных нерестилищах, например, на озере Чеше, насчитывали до 1000 особей[3]. На Северном Кавказе плотность популяции достигает 33,1—179 особей/м³ воды, местами на 1 м прибрежной линии встречается 5—6 особей, хотя общая численность небольшая. В Грузии в некоторых популяциях может насчитываться примерно 6000—7000 взрослых тритонов, которые для размножения используют несколько водоёмов, в каждом из которых встречается от десятка до нескольких сотен особей[13]. В центре ареала в подходящих биотопах обычный, в некоторых местах многочисленный вид, являющийся фоновым, однако популяции на периферии ареала обычно небольшие и изолированные[13][5]. В целом плотность популяции постепенно снижается к востоку и северу[13]. Наиболее многочисленные популяции этого вида размножаются в олиготрофных горных озёрах, в которых полностью отсутствует рыба, таких как Малая Рица, Чеше, Дабадзвели и Кахиси, в окрестностях которых они обитают[15]. Общая численность всех популяций малоазиатского тритона невелика из-за малого количества подходящих для размножения водоёмов[5].
Естественные колебания численности тритонов могут происходить из-за изменения гидрологического и гидрохимического режимов водоёмов: заиливания, заболачивания, резкого снижения уровня воды, пересыхания, возрастания эвтрофикации воды и т. д.[3][21] К массовой гибели этих животных, по предположению исследователей, могут приводить также периодические сильные похолодания в течение зимы, приводящие к вымерзанию тритонов на зимовках[22].
Образ жизни
[править | править код]Зимуют тритоны обычно на суше в трухлявых деревьях, пустотах под камнями, в норах. Однако в Восточной Грузии зимовка нередко происходит в небольших родниках в лесу, а на Западном Кавказе взрослые особи регулярно зимуют в воде. К местам зимовок приходят в сентябре — октябре, выходят из зимних убежищ на низких высотах в феврале — марте, на высокогорьях в апреле — мае. В Краснодарском крае наблюдалось появление в водоёмах в конце ноября — январе. Возможно, здесь зимовка происходит не всегда. Выход из зимовки происходит при подъёме средней температуры воздуха примерно до +5…+15 °C, почвы до +1 °C и воды +5…+9 °C[23]. Основным стимулом к перемещению в водоёмы для размножения (репродуктивной миграции), в частности на Западном Кавказе, являются атмосферные осадки в виде дождей, моросящих там в начале апреля. Именно после дождей тритоны появляются в водоёмах в больших количествах. Первыми к местам размножения, при температуре воды +3°…+5° С, приходят преимущественно самцы (70—80 % особей), самки появляются в водоёмах позже[24][23]. В водоёмах самцы ведут преимущественно нектонный образ жизни, проводя значительную часть времени в толще воды на водной растительности, чему способствует сильное развитие у них в это время плавниковых складок. В то же время самки чаще передвигаются по дну, меньшее развитие у них плавниковых складок снижает возможности их плавания, они менее активны и более скрытны[25][26]. В водоёмах тритоны активны в светлое время суток, на суше — в тёмное время, прячась днём под камнями, корой гнилых пней, упавшими деревьями, во мху, в пустотах грунта и т. д., откуда в дневное время могут появляться лишь в пасмурную погоду[5].
Сразу после окончания размножения взрослые тритоны покидают водоёмы: в предгорных районах в конце апреля — июне, на средних высотах в конце июня — июле, на высокогорьях в конце августа — начале сентября[5]. Самки покидают водоёмы в среднем на две недели позже самцов[5][3]. В целом взрослые тритоны проводят в воде 4—5 месяцев[5]. Выход тритонов на сушу после икрометания связан в основном с повышением температуры воды. Если температура не поднимается выше +18…+20 °C, они могут задерживаться в водоёмах на продолжительное время[24]. Молодые тритоны после выхода на сушу могут удаляться от водоёмов на расстояния до по меньшей мере 4—5 км. Взрослые особи, как правило, держатся вблизи водоёмов, но иногда могут тоже удаляться от них на такие же расстояния[5]. Когда на Черноморском побережье Кавказа выдаются тёплые зимы, тритон здесь не зимует и появляется в водоёмах в конце ноября — декабре. В холодные зимы, когда температура воздуха по ночам понижается до −10° С и ниже, тритоны выходят из зимовки на побережье в феврале, а в высокогорьях в мае[5][3]. На востоке ареала тритон появляется в водоёмах в конце марта — апреле[5].
В субсредиземноморских ландшафтах с биотопами, имеющими повышенную сухость, тритоны могут оставаться в этих биотопах всё лето, перемещаясь к речным истокам с холодной насыщенной кислородом водой[3].
Питание
[править | править код]После перехода к активному питанию личинки питаются мелкими ракообразными. Позже их рацион расширяется, они начинают поедать моллюсков и насекомых. В целом характер избирательности питания у малоазиатского тритона сходен с таковым у гладких тритонов (Lissotriton), однако его личинки поедают больше бентоса[27]. Так, на ранних стадиях развития, когда личинки только начинают активно питаться, почти 80 % их рациона составляют жаброногие ракообразные из семейств хидорид[28] (например, рачок Chydorus sphaericus) и дафниевых (виды рода Simocephalus[29]). В значительно меньших количествах они поедают также ракушковых рачков из семейства Cyprididae, циклопов, дафниевых из рода Daphnia и моинид (виды рода Moina) и совсем немного брюхоногих моллюсков (в частности, из семейства катушек)[28][29]. На поздних стадиях, при формировании задних конечностей, доля хидорид в рационе несколько уменьшается, до 75 %, немного меньше поедают они и ракушковых. Зато возрастает количество потребления циклопов и дафний, начинают поедать бентосных личинок комаров-звонцов. В самом конце личиночного развития, перед метаморфозом, количество хидорид в рационе тритонов уже едва превышает половину, ещё больше возрастает поедание циклопов, поедают ракушковых и дафний[28]. Интенсивность питания личинок выше в ночное время, чем днём, и наиболее высока около 3 часов ночи, когда температура снижается до примерно +14 °С. Наименее интенсивно личинки питаются в вечернее время, около 18 часов, когда температура повышается до приблизительно +22 °С. В зависимости от времени суток изменяется также и рацион личинок: в ночное время в нём повышается доля дафниевых ракообразных (с примерно 3 % от общего количества экземпляров добычи в 18 часов до около 27 % в 3 часа), днём несколько увеличивается количество брюхоногих моллюсков (с 10 % в 0 часов до 26 % в 18 часов)[30]. В целом на протяжении всего развития в рационе личинок преобладают как по количеству экземпляров, так и по биомассе мелкие ракообразные[25].
В период метаморфоза интенсивность питания личинок и разнообразие их пищи резко снижаются. Сеголетки сразу после метаморфоза питаются только сухопутной добычей: коллемболами (их количество в рационе может достигать почти половины от общего числа экземпляров добычи и 20 % общей биомассы[25]), личинками двукрылых насекомых, жуков водолюбов и листоедов, тлями. Взрослые тритоны в водоёмах поедают в основном брюхоногих моллюсков (виды семейства катушек, от трети до половины рациона), ракообразных (в первую очередь ракушковых, а также бокоплавов и водяных осликов[5]) и насекомых (личинок мокрецов, комаров и других). При этом гаммарид и равноногих ракообразных они, по-видимому, поедают чаще, чем другие виды тритонов Кавказа. Это может быть связано с тем, что часть особей размножается в заводях ручьёв, где в основном обитают эти ракообразные, тогда как другие виды тритонов откладывают икру только в полностью стоячих водоёмах. В водной фазе жизни взрослые малоазиатские тритоны нередко поедают других земноводных: личинок тритона Ланца, головастиков кавказской крестовки, малоазиатской и озёрной лягушек, а также икру восточной квакши[27] (её едят самки, у которых она может составлять до половины рациона в это время[31]) и тритона Ланца[32]. Иногда они поедают попадающих в воду сухопутных беспозвоночных. В целом взрослые тритоны в воде чаще всего захватывают добычу размером 0,6—4,0 мм. При этом самки питаются интенсивнее самцов, поедая больше пищи по её относительной массе, что, как предполагается, связано с их более высокими энергетическими затратами на размножение. Кроме того, самки поедают больше бентосных форм, поскольку они менее активны и более скрытны и держатся преимущественно у дна[25]. В наземный период жизни взрослые тритоны питаются червями, мокрицами, наземными насекомыми (например, жуками долгоносиками и двукрылыми) и моллюсками, в небольших количествах могут поедать двупарноногих многоножек из группы Juliformia[9][25].
Размножение
[править | править код]
Малоазиатские тритоны размножаются в зависимости от погоды (суровости зимы) и высоты местности обитания в конце февраля — июле[28]: в предгорьях — в феврале — марте, на средних высотах — в апреле — мае, на высокогорье — в июне — июле[3]. В зависимости от суровости и продолжительности зимы эти сроки могут сдвигаться к более ранним или более поздним почти на месяц[10]. Тем не менее в большинстве мест пик размножения приходится на апрель — май[28]. Период размножения продолжителен даже в пределах одной популяции[33] и может длиться от 1 до 2 месяцев[10]. Такая растянутость периода размножения приводит к тому, что в одних и тех же водоёмах одновременно могут находится и икра, и личинки на последних стадиях развития, у которых вскорости должен начаться метаморфоз[5].
Размножаются тритоны в озёрах, прудах, болотах, канавах, больших лужах и бочажках ручьёв, карстовых ямах-воронках, расположенных как в густых лесах, так и на полностью открытых местах. Как правило, в этих водоёмах много погружённой травянистой растительности, которая иногда бывает разрежённая. Для размножения подходят также водоёмы, в которых хотя и мало водной растительности, но присутствует хороший слой прошлогодних опавших листьев на дне, служащих тритонам убежищем и местом для откладывания икры. Площадь таких водоёмов варьирует от 1 или нескольких квадратных метров до 400, а по другим данным, даже до примерно 1000 м², глубина в пределах от 10 см до 2—3 м, pH = 5,6—7. Реже малоазиатские тритоны встречаются в небольших проточных ручьях с глинистым дном, в которых обязательно присутствуют ямы[13][3]. При возникновении запруд или искусственных водоёмов малоазиатские тритоны их быстро заселяют[5].
В размножении участвуют только крупные особи: самцы с общей длиной тела от 12,6 см, самки — от 10,4 см[3]. В брачный период самцы проявляют территориальное поведение и перед размножением устраивают брачные турниры. Каждый из участвующих в размножении самцов занимает определённую часть водоёма, площадь которой может составлять от 1 до 4,2 м², и защищает свою индивидуальную территорию от других самцов, агрессивно встречая каждого вторгшегося на неё соперника. При этом тритоны принимают демонстрационные позы и кусают противников[28][10]. Заметив нарушителя границ своего участка, самец обычно резким рывком направляется к нему. В 1—2 см от соперника он останавливается, зависая в воде, при малейшем движении противника он нападает на него, впиваясь в основание его гребня сбоку или в переднюю лапу, резко дёргая её и сотрясаясь всем телом. Некоторые такие атаки могут длиться до 5—7 минут, пока нарушитель не вырвется и не покинет территорию. Хозяин при этом продолжает преследование и может неоднократно повторять нападения. В таких брачных турнирах более мелкие самцы могут яростно атаковать и в результате побеждать более крупных самцов. Иногда на приграничных территориях своих брачных участков самцы могут взаимно атаковать друг друга. В таких случаях агрессию проявляют оба самца, они свиваются в клубок, заглатывая лапы друг друга, совершают резкие рывки и часто бьют хвостами. В некоторых случаях такие драки заканчиваются гибелью одного из обессилевших соперников. После гибели соперника самец продолжает ухаживать исключительно за своей самкой, не проявляя интереса к другой. Предполагается, что территориальное поведение и брачные турниры самцов являются фактором, определяющим плотность популяции тритонов в водоёме и способствующим отбору[10].
Перед размножением самец совершает брачный танец. Вначале самец, приблизившись к самке, поворачивается под углом 30—45° ко дну водоёма и демонстрирует ей один бок своего тела, совершая при этом резкие движения туловищем и хвостом перед её мордой. Иногда самец встает на переднюю и заднюю конечности правой либо левой стороны тела, совершая при этом волнообразные движения хвостом перед самкой и у неё над головой. В конце танца самец, двигаясь, изгибает хвост и останавливается, а самка приближается к нему и касается его изогнутого хвоста[28]. После этого самец откладывает сперматофор грушевидной формы и длиной около 1 см[10], который самка захватывает своей клоакой, откуда он переносится в сперматеку. Сперматозоиды из сперматеки оплодотворяют икринки при их откладывании[28]. Откладывание сперматофоров самцами происходит при температуре воды +5…+7 °С. После завершения икрометания самец теряет интерес к прежней самке и начинает ухаживание за другой[10].
Самки начинают икрометание, когда температура воды достигнет +7…+9 °С[10]. Во время икрометания самец не покидает самку. Перед откладыванием икры самка долго, ползая, обследует дно водоёма в поисках подходящего места. Обычно она останавливается в наиболее мелководной, хорошо прогреваемой части водоёма и там зарывается в листву. Непосредственно перед откладыванием икры самка выходит из своего укрытия, и самец стимулирует её к размножению: он подплывает и разворачивается к ней мордой на уровне брюха, затем, растопырив лапы, повисает на боку спиной к самке. Самец совершает частые колебания хвостом и гребнем, после чего наносит в брюхо самки несколько ударов для стимулирования начала икрометания. Такое ухаживание длится 2—3 минуты. После этого около половины минуты пара отдыхает, и процесс повторяется снова несколько раз[10]. Самка откладывает икринки каждую отдельно или короткими цепочками в прибрежной полосе водоёмов на глубине от 5 до 50 см (обычно 15—20 см), прикрепляя задними лапками к водным растениям, на злаки (чаще отмершие) и мелколистные растения, иногда на упавшие в воду ветки и листья или просто на дно[34][35][10][5]. Икра откладывается порциями, по 9—75 икринок за один раз[34][10]. Иногда самка прерывает икрометание, скрываясь от самца. Через несколько минут, а иногда часов, брачные игры возобновляются. Самка откладывает икру в течение 10—12 суток[10]. Всего за сезон самка в природе откладывает от 59 до 240 икринок[34], в среднем около сотни[5]. При разведении в неволе с применением гормональной стимуляции созревания гонад одна самка за сезон может отложить до 803 икринок, откладывая их в течение 11—69 суток, до 28 икринок в сутки[36]. Икринки коричневатого цвета[37], овальной формы[36]. Размер икринок со студенистой оболочкой от 2,0—2,6 × 3,1—3,6 мм[36] до 3 × 4 мм, без оболочки — 1,8 × 2,3 мм[2]. Икра малоазиатского тритона отличается от икры размножающегося в тех же водоёмах тритона Ланца в первую очередь размером икринок, поскольку икринки первого превосходят икринки второго по размеру не менее чем в 1,5 раза[38]. В брачный период тритоны интенсивно питаются бокоплавами, пиявками и другими водными беспозвоночными и головастиками и чаще, чем в другое время, всплывают к поверхности воды за очередной порцией воздуха[10].
Развитие
[править | править код]Развитие эмбриона в икринке в природе длится от 12 до 30 суток[34]. Очень высокая смертность эмбрионов в икре, предличинки в среднем выклёвываются только из 14 % икринок, а некоторые кладки погибают полностью[5]. При искусственной инкубации при температуре +19,0…+23,5 °C предличинки выклёвывались из икринок через 8—20 суток (в среднем через 14,5). При этом они выклёвывались из 50—79 % всех отложенных икринок[36]. Общая длина предличинки сразу после вылупления из икринки составляет 7—8 мм[34], при разведении в неволе вылуплялись предличинки длиной до 10,5 мм[36]. В водоёмах на высоте 600—1000 м выклев предличинок происходит в конце апреля — начале мая, а на высоте 1300—1400 м в конце мая — начале июля[5].
Плотность личинок может варьировать от 4,6 до 47 особей на 10 л воды, максимального значения она достигает сразу после пика выклева[39]. При выращивании в неволе первые личинки при температуре +18…+23 °C переходят к активному питанию (поедают мелких беспозвоночных) через 12—25 суток после откладки икринок, в среднем через 20,8 дня, при общей длине тела 10,3—10,9 мм[36] и массе около 3,8 мг[25]. Во время перехода к активному питанию личинки малоактивны, держатся в основном среди водной растительности недалеко от мест выклева, интенсивно питаются только мелкими рачками. В дальнейшем личинки становятся более активными, расселяясь шире по водоёму, начинают поедать также личинок насекомых и моллюсков[25].
У малоазиатского тритона личинки лимнофильного (прудового) типа: имеют крепкое телосложение, длинные жаберные ветви и лепестки, высокие хвостовые плавниковые складки[40], спинная хвостовая складка начинается сразу за головой, на уровне жаберных ветвей, на передних конечностях по четыре пальца, на задних по пять, пальцы относительно короткие, без коготков, конец хвоста заострённый[41]. На боках и спине личинки имеют неправильной формы частично сливающиеся светлые пятна, хвост пигментирован почти равномерно. У старших личинок имеются светлые пятна за глазами и чёткая граница между тёмными боками и светлым брюхом. Рисунок на спине образован единичными меланофорами и их небольшими скоплениями. Продольный диаметр глаза меньше расстояния между ноздрями[42]. Внешне личинка малоазиатского тритона похожа на личинку тритона Ланца[2].
Личинки держатся в основном на дне и среди водной растительности[40], появляясь на свободных от растительности участках водоёмов только в конце своего развития[43]. Скорость роста личинок может составлять от 0,07 до 0,34 мм/сутки[44] и зависит в первую очередь, помимо наличия корма, от температуры и объёма воды[45]. При температуре +17,5…+18,5 °C личиночное развитие (от начала питания мелкими беспозвоночными до выхода метаморфизировавшего животного на сушу) может продолжаться от 69 до 157 суток[46]. При повышении температуры до +23…+27 °C личинки развиваются интенсивно, но при этом проходят очередные стадии развития, имея более мелкие размеры[47]. В любом случае личинки растут, увеличиваясь в размерах, вплоть до начала метаморфоза, признаком которого является уменьшение жабр. При прохождении метаморфоза молодые тритоны несколько уменьшаются, в результате сразу после выхода на сушу они могут быть немного меньше, чем были на последней личиночной стадии в воде[48]. Перед метаморфозом длина тела личинок может достигать 5,1 см, хвоста — 5,0 см[5], а масса по сравнению со стадией перехода на активное питание увеличивается в 49,5 раз[25].
Метаморфоз проходит на западе ареала в июле — августе, на востоке — в конце сентября — октябре[5]. В предгорьях сеголетки покидают водоёмы в августе, на высокогорье — в конце сентября[3]. Таким образом, весь период развития от откладки икринок до выхода на сушу сеголеток обычно длится 5,5—6 месяцев[5]. В отдельных горных прудах личинки, которые вылупились из поздних кладок, перезимовывают и завершают превращение и выходят на сушу только на следующий год[28]. Растянутость периода выхода метаморфов зависит от растянутости икрометания и связана со временем откладки икры[3]. Молодые тритоны сразу после метаморфоза имеют общую длину тела 2,2—4,4 см[2]. Прошедшие метаморфоз особи выходят на сушу и не возвращаются в водоёмы до достижения половой зрелости[49].
Половая зрелость наступает в возрасте от 2—3 лет у тритонов из популяций, обитающих на малых высотах, до 6 лет у высокогорных[50] при общей длине самцов около 12,6 см и самок 10,4 см[5]. Соотношение полов в популяциях примерно равное[5]. Продолжительность жизни в природе составляет 8—12 лет, максимальная, изредка — до 21 года[28].
В озере Малая Рица в Абхазии в 2007 году среди многочисленных нормальных взрослых особей была найдена педоморфная самка малоазиатского тритона. Это был первый случай обнаружения педоморфной формы этого вида амфибий. Самка имела общую длину тела 10,4 см при длине туловища с головой 5,5 см, то есть достигала практически тех же размеров, что и метаморфизировавшие особи. Она имела хорошо развитые наружные жабры, ровный спинной плавник и несколько более светлую окраску[51].
Враги
[править | править код]Икру малоазиатского тритона могут поедать самки своего же вида и тритоны Ланца; личинок — пресноводный краб Potamon potamios, водяной жук плавунец окаймлённый, интродуцированный обыкновенный карась, малоазиатская и озёрная лягушки, кавказская крестовка и болотная черепаха, сеголеток может массово поедать озёрная лягушка, на взрослых тритонов нападают обыкновенный, водяной и колхидский ужи, врановые птицы (чёрная ворона, ворон и сойка) и водяная кутора[52][3]. В последнее время основным врагом тритона стал завезённый человеком американский енот-полоскун, который на нерестилищах может уничтожать от 50 до 100 % половозрелых особей[3]. Каннибализм в местах размножения в форме оофагии, по-видимому, широко распространён, особенно среди самок, которые чаще самцов контактируют с отложенной икрой. В неволе наблюдались случаи каннибализма среди личинок малоазиатского тритона[20].
У малоазиатского тритона отмечены 3 вида гельминтов: плоский червь Brachycoelium salamandrae из подкласса дигенетических сосальщиков и круглые черви Megalobatrachonema terdentatum и Neoxysomatium caucasicum из подотряда Ascaridina. Икринки иногда поражает сапролегниевый оомицет[20], хотя икра малоазиатского тритона менее подвержена гибели из-за этого, чем икра тритонов Ланца и Карелина. Предполагается, что это может быть одной из причин относительно высокого по сравнению с ними процента выхода предличинок этого вида из икры[53].
У содержащихся в неволе малоазиатских тритонов был обнаружен патогенный, вызывающий хитридиомикоз хитридиевый грибок Batrachochytrium salamandrivorans[54].
Классификация
[править | править код]
Малоазиатский тритон был впервые научно описан немецким зоологом Арнольдом Бертольдом в 1846 году под названием Triton ophryticus по экземпляру, собранному в Грузии в районе Тифлиса (типовая территория). Большую часть XX века малоазиатского тритона относили к роду Triturus, выделяя в качестве подвида Triturus vittatus ophryticus в составе вида Triturus vittatus. В начале XXI века на основании результатов генетических исследований род Triturus был разделён на несколько, в том числе был выделен род Ommatotriton с единственным видом Ommatotriton vittatus. В 2005 году уже этот вид был разделён на два — Ommatotriton vittatus и Ommatotriton ophryticus, каждый из которых, в свою очередь, разделяли на подвиды. Данное разделение было проведено на основании уровня дивергенции прежних подвидов вида vittatus, который соответствовал тому, что был определён у видов тритонов из других родов[2]. В 2005 году у вида Triturus ophryticus был описан подвид Triturus ophryticus nesterovi Litvinchuk, Zuiderwijk, Borkin et Rosanov, 2005, обитающий на северо-западе Турции от региона Мраморного моря до окрестностей Акчая в северной части Малой Азии[12]. В 2013 году этот подвид был выделен в самостоятельный вид Ommatotriton nesterovi (Litvinchuk, Zuiderwijk, Borkin et Rosanov, 2005)[55]. Ареалы Ommatotriton ophryticus и Ommatotriton nesterovi вплотную соприкасаются между собой, граница между ними находится на черноморском побережье Турции и проходит вдоль 36,5° восточной долготы. В этом месте эти два вида образуют межвидовые гибриды[56].
Таким образом, в настоящее время Ommatotriton ophryticus является одним из трёх видов монофилетического рода малоазиатских, или полосатых тритонов (Ommatotriton), распространённого в Юго-Восточной Европе и Юго-Западной Азии в Восточном и Южном Причерноморье, на Кавказе и в Восточном Средиземноморье от Краснодарского края на севере до Израиля на юге и от южного побережья Мраморного моря на западе до западных склонов Эльбруса на востоке[11]. Скорее всего, наиболее близкородственным видом малоазиатскому тритону является Ommatotriton nesterovi, однако у исследователей пока нет единого мнения по этому поводу[57].
Происхождение
[править | править код]Хотя обнаружить ископаемые остатки малоазиатского тритона до сих пор не удалось, исследователи, на основании изучения аллозимов[англ.] этих амфибий и палеогеографических реконструкций, предполагают, что в раннем и среднем олигоцене (38—35 млн лет назад) предковая популяция этого вида населяла Западную Европу. В позднем олигоцене (около 23 млн лет назад) Балкано-Малоазиатский полуостров был соединён сушей с Центральной Европой. В это время предковая популяция малоазиатского тритона могла проникнуть на этот полуостров. В раннем миоцене (около 20 млн лет назад) он отделился от Центральной Европы (на месте их соединения образовался пролив), в то же время соединившись с противоположной стороны с Западной Азией. Около 17 млн лет назад перешеек между Малоазиатским и Аравийским массивами суши также исчез. Это разделение могло быть причиной расхождения рода Ommatotriton (на сирийско-израильском средиземноморском побережье) с другими родами тритонов. В позднем сармате (около 9,5 млн лет назад) пролив исчез, и предки Ommatotriton ophryticus расселились на территории, которая сейчас является южным побережьем Чёрного моря. Согласно результатам изучения аллозимов дивергенция (расхождение видовых признаков) между малоазиатским (Ommatotriton ophryticus) и ближневосточным (Ommatotriton vittatus) тритонами произошла примерно 10,9—7,4 млн лет назад, в позднем миоцене. Предковая популяция вида Ommatotriton nesterovi проникла в западную часть Малой Азии примерно 5,0—7,3 млн лет назад. В это же время, в самом конце миоцена, Ommatotriton ophryticus мог проникнуть на Большой Кавказ, который был связан сушей с Малым Кавказом[12]. Однако, по мнению других учёных, дивергенция малоазиатского и ближневосточного тритонов произошла значительно раньше — около 25 млн лет назад вследствие поднятия горных систем в восточной части Малой Азии. Предполагается также, что во время последнего ледникового максимума ареал малоазиатского тритона претерпел сокращение и фрагментацию, однако эти земноводные выжили в многочисленных ледниковых рефугиумах[57].
Основные угрозы
[править | править код]Общая численность малоазиатского тритона невелика и снижается[3][58]. Помимо хищничества интродуцированного енота-полоскуна, причинами сокращения популяции являются также браконьерский вылов тритонов и уничтожение мест обитания, которые, особенно на побережье, в последние десятилетия подверглись резкому изменению в результате строительства курортной инфраструктуры и возрастания рекреационной нагрузки[3]. Яркая внешность самцов в брачном наряде делает их желанным объектом содержания для любителей террариумов, в связи с чем в 1990-х — начале 2000-х годов имел место массовый нелегальный вылов этих тритонов в природе для продажи. Малоазиатский тритон регулярно и в больших количествах продавался в Ереване, Киеве, Москве, Сочи, Тбилиси и т. д. Временами тритонов продают и сейчас[20]. К сокращению популяции малоазиатского тритона приводят также вырубка лесов, мелиорация, приводящая к уничтожению мелких водоёмов, перевыпас скота (связанное с этим загрязнение скотом водоёмов), урбанизация, промышленное загрязнение, использование пестицидов и искусственных удобрений на сельскохозяйственных угодьях, прокладка дорог и движение автотранспорта[20][21], разрушение, реконструкция, загрязнение и зарыбление (например, интродукция обыкновенной гамбузии на Пицундской низменности) водоёмов, где эти тритоны размножаются[15]. Во многих местах осушение водоёмов уже привело к полному исчезновению этих амфибий. Прогнозируемые учёными глобальные изменения климатических условий (глобальное потепление) в ближайшие полвека могут привести к усилению аридизации ландшафтов на Кавказе, что приведёт к исчезновению пригодных для этого вида биотопов во многих местах его обитания[3], в первую очередь на севере и юге ареала и, особенно, на территории Восточной Грузии и Армении[15]. В то же время создание искусственных прудов ведёт к повышению численности малоазиатского тритона[59].
Охрана
[править | править код]
В природоохранном отношении малоазиатский тритон находится в наилучшем положении среди хвостатых земноводных Кавказа. Однако в связи с узким ареалом его в целом считают редким видом[20]. Международным союзом охраны природы и природных ресурсов малоазиатскому тритону присвоен охранный статус вида, близкого к уязвимому положению[58]. Малоазиатский тритон был занесён в Красную книгу СССР как редкий вид с сокращающейся численностью[60]. В настоящее время как сокращающийся в численности и/или распространении уязвимый вид малоазиатский тритон занесён в Красную книгу России[3]. Как близкий к уязвимому положению вид занесён в Красную книгу Грузии[20][61], как находящийся на грани полного исчезновения — в Красную книгу Армении[14], как уязвимый, резко сократившийся в численности за последнее десятилетие реликтовый эндемичный вид, имеющий ограниченное число мест обитания, внесён в Красные книги Краснодарского края[4] и Республики Адыгея[62], как редкий, спорадически распространённый реликтовый вид — в Красную книгу Карачаево-Черкесской Республики[63]. Занесён в Приложение ІІІ Бернской конвенции[5].
Обитает среди прочего на нескольких природоохранных территориях: охраняется в Кавказском биосферном заповеднике, где встречается спорадично по всей территории кроме высокогорий (выше 1900 м) Главного Кавказского хребта восточнее горы Чугуш[6], Лиахвском заповеднике[64] и Сочинском национальном парке[65], встречается на Архызском участке Тебердинского национального парка (в горных районах долины Архыза и, по некоторым данным, в озере в долине реки Кизгыч)[66], в национальном парке Борчка-Карагёль на северо-востоке Турции[67], формально охраняется в нескольких заказниках: Псебайском, Туапсинском, Горячеключевском и других[3]. В 1986 году[16] и середине 1990-х годов сотрудниками Московского зоопарка были проведены успешные попытки разведения малоазиатского тритона в неволе и реинтродукции его сеголеток в дикую природу в Кавказском заповеднике[3][20]. Успешно была проведена его реинтродукция в Армении[59]. В качестве необходимых дополнительных мер охраны рекомендуется полное уничтожение в местах обитания вида завезённого енота-полоскуна, а также создание трансграничной (российско-грузинской) природоохранной территории «Северная Колхида», объединяющей Кавказский биосферный заповедник, Сочинский национальный парк и Рицинский реликтовый национальный парк, общей площадью более 547 000 га[3]. Для охраны вида важно, чтобы все горные озёра, где обитают его наиболее многочисленные популяции, были включены в состав природоохранных территорий. Способствовать увеличению численности вида в местах его обитания могло бы также создание новых прудов[15]. Кроме того, необходимы выявление нерестилищ тритонов и организация в этих местах микрозаповедников или памятников природы, запрещение отлова этих земноводных в коммерческих и учебных целях и ограничение их научного отлова[4].
Примечания
[править | править код]- ↑ 1 2 Frost D.[англ.]. Ommatotriton ophryticus (Berthold, 1846). Amphibian Species of the World 6.2, an Online Reference. American Museum of Natural History. Дата обращения: 20 сентября 2023. Архивировано 29 января 2023 года.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 Кузьмин, 2012, с. 88.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 Туниев Б. С. Малоазиатский тритон Ommatotriton ophryticus (Berthold, 1846) // Красная книга Российской Федерации. Том «Животные» / под ред. Н. Б. Ананьевой, А. В. Белоусовой, Т. А. Бритаева и др.; председ. гл. ред. колл. Д. С. Павлов. — М.: ФГБУ «ВНИИ Экология», 2021. — С. 412—413. — 1128 с. — ISBN 978-5-6047425-0-1.
- ↑ 1 2 3 4 5 6 7 8 9 10 Туниев Б. С. 374. Тритон малоазиатский Ommatotriton ophryticus (Berthold, 1870) // Красная книга Краснодарского края. Животные. III издание / Отв. ред. А. С. Замотайлов, Ю. В. Лохман, Б. И. Вольфов. — Краснодар: Адм. Краснодар. края, 2017. — С. 479—481. — 720 с. — ISBN 978-5-6040022-5-4.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 Боркин Л. Я. Малоазиатский тритон // Земноводные и пресмыкающиеся (Серия: Энциклопедия природы России) / Ананьева Н. Б., Боркин Л. Я., Даревский И. С., Орлов Н. Л. / ред. серии д. б. н., к. г. н. А. А. Минин. — М.: ABF, 1998. — С. 56—59. — 576 с. — ISBN 5-87484-066-4.
- ↑ 1 2 3 4 5 Иваненко А. М., Ковалёв В. В. Амфибии и рептилии Кавказского заповедника. — Майкоп: НАБУ-Кавказ, 2019. — С. 20—21. — 86 с.
- ↑ Пескова Т. Ю., Плотников Г. К. Сравнительная морфометрия малоазиатского тритона из различных популяций Северо-Западного Кавказа // Материалы Международной научной конференции «Биосфера и человек» (Адыгейский государственный университет, 24—25 октября 2019 года) / науч. ред. А. В. Шаханова. — Майкоп: ЭлИТ, 2019. — С. 88—90.
- ↑ 1 2 3 Кузьмин, 2012, с. 31, 88.
- ↑ 1 2 Дунаев Е. А., Орлова В. Ф. Земноводные и пресмыкающиеся России: атлас-определитель. — 2-е изд. — М.: Фитон XXI, 2017. — С. 24. — 328 с. — ISBN 978-5-906811-32-5. Архивировано 31 октября 2023 года.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Туниев Б. С. К размножению малоазиатского тритона // Вестник зоологии[укр.]. — 1982. — № 2. — С. 69—70.
- ↑ 1 2 Üzüm N., Avcı A., Olgun K., Bülbül U., Fahrbach M., Litvinchuk S. N., Wielstra B. Cracking cryptic species: external characters to distinguish two recently recognized banded newt species (Ommatotriton ophryticus and O. nesterovi) (англ.) // Salamandra. — 2019. — Vol. 55, iss. 2. — P. 131—134.
- ↑ 1 2 3 Litvinchuk S. N., Zuiderwijk A., Borkin L. J., Rosanov J. M. Taxonomic status of Triturus vittatus (Amphibia: Salamandridae) in western Turkey: trunk vertebrae count, genome size and allozyme data (англ.) // Amphibia-Reptilia. — 2005. — Vol. 26. — P. 305—323.
- ↑ 1 2 3 4 5 6 7 8 9 10 Кузьмин, 2012, с. 89.
- ↑ 1 2 Kalashian M., Aghababyan K., Zarikian N., Gabrielyan B., Arakelyan M., Ghazaryan A. Fauna of Armenia // Biodiversity of Armenia / ed. by G. Fayvush. — Cham: Springer, Cham, 2023. — P. 252. — 440 p. — ISBN 978-3-031-34331-5.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 Литвинчук С. Н. Распространение и природоохранный статус малоазиатского тритона, Ommatotriton ophryticus (Amphibia: Caudata) // Nature Conservation Research. Заповедная наука. — 2017. — Т. 2, вып. 1. — С. 33—39.
- ↑ 1 2 Кукушкин О. В., Доронин И. В., Туниев Б. С., Ананьева Н. Б., Доронина М. А. Интродукция земноводных и пресмыкающихся на Кавказе и в Крыму: общий обзор и некоторые факты // Современная герпетология. — 2017. — Т. 17, вып. 3/4. — С. 157—197. — doi:10.18500/1814-6090-2017-17-3-4-157-197.
- ↑ Tuniyev B. S., Koval A. G., Vargovitsh R. S. Amphibians and reptiles in the caves of the Greater Caucasus (англ.) // Nature Conservation Research. Заповедная наука. — 2021. — Vol. 6, iss. 1. — P. 1—16. — doi:10.24189/ncr.2021.010. Архивировано 4 октября 2023 года.
- ↑ Кузьмин, 2012, с. 104.
- ↑ Пястолова, Тархнишвили, 1989, с. 132.
- ↑ 1 2 3 4 5 6 7 8 Кузьмин, 2012, с. 91.
- ↑ 1 2 Кузьмин, 2012, с. 269—270.
- ↑ Пястолова, Тархнишвили, 1989, с. 129.
- ↑ 1 2 Кузьмин, 2012, с. 89—90.
- ↑ 1 2 Пястолова, Тархнишвили, 1989, с. 33.
- ↑ 1 2 3 4 5 6 7 8 Кузьмин С. Л., Тархнишвили Д. Н. Возрастная динамика питания симпатрических тритонов Кавказа // Зоологический журнал. — 1987. — Т. 66, вып. 2. — С. 244—258.
- ↑ Пястолова, Тархнишвили, 1989, с. 122.
- ↑ 1 2 Кузьмин, 2012, с. 90—91.
- ↑ 1 2 3 4 5 6 7 8 9 10 Кузьмин, 2012, с. 90.
- ↑ 1 2 Пястолова, Тархнишвили, 1989, с. 95.
- ↑ Пястолова, Тархнишвили, 1989, с. 110.
- ↑ Пястолова, Тархнишвили, 1989, с. 124.
- ↑ Кузьмин, 2012, с. 84.
- ↑ Кузьмин, 2012, с. 261.
- ↑ 1 2 3 4 5 Кузьмин, 2012, с. 88, 90.
- ↑ Пястолова, Тархнишвили, 1989, с. 37.
- ↑ 1 2 3 4 5 6 Кидов А. А., Матушкина К. А. Заметки к репродуктивной биологии малоазиатского тритона, Ommatotriton ophryticus (Berthold, 1846) на северо-восточной периферии ареала // Вестник Чувашского государственного педагогического университета им. И. Я. Яковлева. — 2017. — Вып. 95, № 3—1. — С. 3—9.
- ↑ Кузьмин, 2012, с. 23.
- ↑ Пястолова, Тархнишвили, 1989, с. 27.
- ↑ Пястолова, Тархнишвили, 1989, с. 45.
- ↑ 1 2 Кузьмин, 2012, с. 258.
- ↑ Кузьмин, 2012, с. 25, 26.
- ↑ Кузьмин, 2012, с. 26.
- ↑ Пястолова, Тархнишвили, 1989, с. 44.
- ↑ Пястолова, Тархнишвили, 1989, с. 88.
- ↑ Пястолова, Тархнишвили, 1989, с. 134.
- ↑ Пястолова, Тархнишвили, 1989, с. 62.
- ↑ Пястолова, Тархнишвили, 1989, с. 61.
- ↑ Пястолова, Тархнишвили, 1989, с. 65, 68.
- ↑ Пястолова, Тархнишвили, 1989, с. 16.
- ↑ Gümüş Özcan Ç., Üzüm N. Body size and age in three populations of the northern banded newt Ommatotriton ophryticus (Berthold, 1846) from Turkey (англ.) // Turkish Journal of Zoology. — 2015. — Vol. 39. — P. 672—679. — doi:10.3906/zoo-1406-35.
- ↑ Skorinov D. V., Novikov O., Borkin L. J., Litvinchuk S. N. Two new cases of paedomorphosis in the Caucasian newts: Ommatotriton ophryticus (the first record) and Lissotriton vulgaris lantzi (англ.) // Russian Journal of Herpetology. — 2009. — Vol. 16, iss. 1. — P. 16—18.
- ↑ Кузьмин, 2012, с. 91, 232.
- ↑ Пястолова, Тархнишвили, 1989, с. 48.
- ↑ Fitzpatrick L. D., Pasmans F., Martel A., Cunningham A. A. Epidemiological tracing of Batrachochytrium salamandrivorans identifies widespread infection and associated mortalities in private amphibian collections (англ.) // Scientific Reports[англ.]. — 2018. — Vol. 8, no. Art. 13845 (2018). — doi:10.1038/s41598-018-31800-z.
- ↑ Bülbül U., Kutrup B. Morphological and genetic variations of Ommatotriton in Turkey (англ.) // Animal Biology[англ.]. — 2013. — Vol. 63, iss. 3. — P. 297—312.
- ↑ Kalaentzis K., Arntzen J. W., Avcı A., van den Berg V., Beukema W., France J., Olgun K., van Riemsdijk I., Üzüm N., de Visser M. C., Wielstra B. Hybrid zone analysis confirms cryptic species of banded newt and does not support competitive displacement since secondary contact (англ.) // Ecology and Evolution[англ.]. — 2023. — Vol. 13, iss. 9. — P. e10442. — doi:10.1002/ece3.10442.
- ↑ 1 2 Van Riemsdijk I., Arntzen J. W., Bogaerts S., Franzen M., Litvinchuk S. N., Olgun K., Wielstra B. The Near East as a cradle of biodiversity: A phylogeography of banded newts (genus Ommatotriton) reveals extensive inter- and intraspecific genetic differentiation (англ.) // Molecular Phylogenetics and Evolution. — 2017. — Vol. 114. — P. 73—81. — doi:10.1016/j.ympev.2017.05.028. Архивировано 19 декабря 2023 года.
- ↑ 1 2 Olgun et al., 2009.
- ↑ 1 2 Кузьмин, 2012, с. 257.
- ↑ Белова З. В. Малоазиатский тритон Triturus vittatus Jenyns, 1835 // Красная книга СССР: Редкие и находящиеся под угрозой исчезновения виды животных и растений. Том 1 / Главная ред. коллегия: А. М. Бородин, А. Г. Банников, В. Е. Соколов и др. — 2-е изд. — М.: Лесная промышленность, 1984. — С. 175—176. — 392 с.
- ↑ Кузьмин, 2012, с. 276—278.
- ↑ Туниев Б. С., Туниев С. Б. 366. Малоазиатский тритон Ommatotriton ophryticus (Berthold, 1846) // Красная книга Республики Адыгея: Редкие и находящиеся под угрозой исчезновения объекты животного и растительного мира. Часть 2: Животные / отв. ред. А. С. Замотайлов; научн. ред.: А. С. Замотайлов, В. И. Щуров, М. И. Шаповалов, Р. А. Мнацеканов. — 2-е изд. — Майкоп: Качество, 2012. — С. 229—230. — 376 с. — ISBN 978-5-9703-0347-4.
- ↑ Доронин И. В. Малоазиатский тритон Ommatotriton ophryticus (Berthold, 1846) // Красная книга Карачаево-Черкесской Республики / Ред. колл.: Кочкаров Х. Б., Текеев Д. К., Зернов А. С., Батчаев Р. И., председ. Онипченко В. Г. — Черкесск: Нартиздат, 2013. — С. 73. — 360 с. — ISBN 978-5-906148-05-6.
- ↑ Туниев Б. С., Лотиев К. Ю., Туниев С. Б., Габаев В. Н., Кидов А. А. Амфибии и рептилии Южной Осетии // Nature Conservation Research. Заповедная наука. — 2017. — Т. 2, вып. 2. — С. 1—23. — doi:10.24189/ncr.2017.002.
- ↑ Туниев Б. С., Рыбак Е. А., Тимухин И. Н., Алиев Х. У. Хорологические закономерности в экотоне Восточно-Средиземноморской и Колхидской биогеографических провинций на территории Сочинского национального парка // Экосистемы. — 2022. — Т. 29. — С. 5—26. — ISSN 2414-4738.
- ↑ Поливанов В. М., Морозова О. В. Земноводные и пресмыкающиеся // Позвоночные животные Тебердинского заповедника (Серия: Флора и фауна заповедников. Вып. 100) / Под ред. И. С. Даревского. — М.: ИПЭЭ РАН, 2002. — С. 8—9. — 60 с.
- ↑ Afsar M., Afsar B., Ayaz D., Çiçek K., Tok C. V. Amphibians of the Borçka-Karagöl Nature Park (Artvin/Turkey) and Vicinity (англ.) // Iğdır University Journal of the Institute of Science and Technology. — 2018. — Vol. 8, iss. 2. — P. 49—55. — doi:10.21597/jist.427880.
Литература
[править | править код]- Кузьмин С. Л. Земноводные бывшего СССР. — 2-е изд. — М.: Товарищество научных изданий КМК, 2012. — С. 88—91. — 370 с. — ISBN 978-5-87317-871-1.
- Пястолова О. А., Тархнишвили Д. Н. Экология онтогенеза хвостатых амфибий и проблема сосуществования близких видов / Отв. ред. акад. В. Н. Большаков. — Свердловск: УрО АН СССР, 1989. — 156 с.
Ссылки
[править | править код]- Olgun K., Arntzen J., Kuzmin S., Papenfuss T., Ugurtas I. H., Tarkhnishvili D., Sparreboom M., Anderson S., Tuniyev B., Ananjeva N. B., Kaska Y., Kumlutaş Y., Avci A., Üzüm N., Kaya U. Northern Banded Newt Ommatotriton ophryticus (англ.). The IUCN Red List of Threatened Species. IUCN (2009 (errata version published in 2019)). doi:IUCN.UK.2009-1.RLTS.T136019A155801190.en. Дата обращения: 21 сентября 2023.
Эта статья входит в число избранных статей русскоязычного раздела Википедии. |